

Ranas terrestres en los ecosistemas surandinos de Ecuador IV: Una nueva especie de Pristimantis (Anura: Strabomantidae) de los páramos sur del Parque Nacional Sangay
Terrestrial frogs in the south Andean ecosystems of Ecuador IV: A new species of Pristimantis (Anura: Strabomantidae) of the southern paramos of Sangay National Park
ACI Avances en Ciencias e Ingenierías
Universidad San Francisco de Quito, Ecuador
Recepción: 10 Agosto 2016
Aprobación: 30 Junio 2017
Resumen: Describimos una nueva especie de rana terrestre Pristimantis de los Andes sur de Ecuador. La nueva especie se caracteriza por la presencia de procesos dentígeros del vómer de contorno triangular; tubérculos ulnares aplanados, dispuestos en una hilera a manera de pliegue; y una distintiva coloración ventral de café claro a café oscuro con verrugas pigmentadas de blanco. Su canto presenta una única nota con frecuencia dominante promedio de 4.23 kHz y dos parciales armónicos, y una duración promedio de 207 ms.
Palabras clave: Cañar, Laguna de Culebrillas, Pristimantis caniari sp. nov., Andes sur.
Abstract: We describe a new species of terrestrial frog of the genus Pristimantis, located in the south Andes of Ecuador. The new species is characterized by: presence of dentigerous processes of vomer that are triangular in outline; ulnar tubercles flattened and disposed in a longitudinal, fold-like row; and a distinctive ventral coloration light to dark brown with white warts. We describe the call of an adult male, a single note with an average frequency of 4.23 kHz and two partial harmonics, and an average duration of 207 ms.
Keywords: Cañar, Culebrillas lacuna, Pristimantis caniari sp. nov., south Andes.
INTRODUCCIÓN
El género Pristimantis constituye el grupo de anfibios terrestres más diverso en el Neotrópico [1-3], con 207 y 201 especies formalmente descritas, Colombia y Ecuador son los países con mayor riqueza del género [3-4]. Las recientes descripciones de Pristimantis en el territorio ecuatoriano [5-9], han acrecentado significativamente el número de especies, así, en los últimos dos años se han descrito más de 15 especies nuevas [5-13]. La alta riqueza de ranas terrestres en Ecuador es también acompañada de un alto porcentaje de endemismo (i.e. 54%) [4], la mayoría de especies endémicas se restringen a las estribaciones de los Andes ecuatorianos [4], reflejando a los ecosistemas andinos como hábitats idóneos para las ranas Pristimantis [1,4,14].
Los páramos y estribaciones del Centro Sur del Ecuador han sido consideradas zonas poco exploradas y su estudio durante la última década ha arrojado el descubrimiento de varias especies nuevas [5-8,12-13,15-20], de las cuales tres han sido reportadas en las tierras altas y paramunas de la región (i.e. Pristimantismazar, P. bambu, P. gulacenio) [18-20]. En este artículo describimos una nueva especie de los páramos de la región, cuya morfología externa es similar a las especies del grupo de Pristimantis orcesi (sensu Hedges et al. [2]), además incluimos un análisis de las variables espectrales y temporales de su canto.
Materiales y Métodos
Análisis morfológico: la descripción de la especie sigue el estándar de Lynch y Duellman [1] y el uso de los caracteres diagnósticos hace referencia a las definiciones e ilustraciones propuestas por Duellman y Lehr [21]. Utilizamos la propuesta de Heinicke et al. [22] para la clasificación sistemática de familia y seguimos los grupos putativos de especies propuestos por Hedges et al. [2] y Padial et al. [23]. Los especímenes colectados fueron sacrificados en una solución de benzocaína, fijados en formalina al 10% y preservados en etanol al 70%. El sexo y edad de los especímenes fueron determinados mediante la identificación de características sexuales secundarias (almohadillas nupciales, hendiduras bucales en los machos y tamaño corporal) e inspección directa de las gónadas a través de incisiones dorsolaterales. Las medidas morfométricas fueron tomadas con un calibrador electrónico (precisión ± 0.01 mm, aproximados a 0.1 mm) siguiendo los comentarios de Duellman y Lehr [21]. Los patrones de coloración en vida fueron extraídos de las notas de campo y fotografías a color. Las coordenadas y elevación de la localidad tipo fueron determinadas en base a las coordenadas registradas con un GPS y las notas de campo del colector. En todo el texto se hacen uso de las siguientes abreviaciones para los créditos fotográficos e información asociada: Salomón Ramírez Jaramillo (SRJ), Carolina Reyes-Puig (CRP) y Mario H. Yánez-Muñoz (MYM).
Los especímenes examinados están depositados en el repositorio de Herpetología del Instituto Nacional de Biodiversidad, Quito (DHMECN), Instituto de Ciencias Naturales (ICN) Universidad Nacional de Colombia, y Colección Herpetológica de la Universidad del Valle del Cauca (UVC). Excepto por las series tipo, los siguientes especímenes fueron examinados:
ECUADOR: Pristimantis orcesi: Tungurahua: Parque Nacional Llanganates, 3510m, DHMECN 1820-1823; Napo: Papallacta, 3910m, DHMECN 1857-1866; Napo: Cuyuja, 3591m, DHMECN 2903. Pristimantis ortizi: Carchi: Julio Andrade - Ipueran, 3420m, DHMECN 2277; Carchi: Laguna de Maicas, 3443m, DHMECN 9346; Sucumbíos: La bonita, 2614m, DHMECN 6389-6390; Pristimantis riveti: Chimborazo: Parque Nacional Sangay - Lagunas Negras, 3600m, DHMECN 1849-1851; Azuay: Molleturo, 3600m, DHMECN 3084; Azuay: Gualaceo - Bosque Aguarongo, 3132m, DHMECN 11877-11878. Pristimantis thymelensis: Carchi: Reserva Ecológica El Ángel, 3900m, DHMECN 1867-1890; Carchi: Lagunas del Voladero, 3420m, DHMECN 2415-2419; Napo: Papallacta, 3910m, DHMECN 1856. COLOMBIA: Pristimantis obmutescens:Departamento del Cauca: Páramo de Puracé, alrededores de la Laguna San Rafael, ICN 2087, holotipo. Pristimantis racemus: Departamento del Valle del Cauca, Tenerife, UVC 8131, 8193. Pristimantis simoteriscus: Departamento del Tolima: Municipio de Cajamarca, Páramo de los Valles, SW of Anaime, Anaime-Santa Helena road, ICN 22835, holotipo. Pristimantis simoterus: Departamento del Tolima: Páramo de Letras, páramo-subpáramo, vereda Albania, Municipio de Herveo, ICN 759, hembra, holotipo.
Análisis bioacústico: los cantos se registraron con una grabadora digital Olympus WS- 750, conectada a un sistema modular Sennheiser K6-C, acoplado a un micrófono de cabezal Sennheiser ME 66, tomando la temperatura y humedad con un termómetro ambiental marca Springfield. Para los análisis acústicos se utilizó el programa Adobe Audition 3.0 a una frecuencia de muestreo de 44.1 kHz y 16 “bits” de resolución [24]; para construir el oscilograma y sonograma se utilizó el programa Raven 1.4 [25] a 256 puntos de resolución de la transformación rápida de Fourier (FFT). Las grabaciones fueron obtenidas el 26 junio 2009 a las 19h55 por SRJ a una temperatura de 3,3° C y 91% de humedad relativa correspondiente al ejemplar DHMECN 10341. Las variables analizadas fueron: (1) Frecuencia dominante: frecuencia de mayor energía medida a lo largo de todo el canto; (2) Frecuencias armónicas: frecuencias más altas que son múltiplo de la frecuencia fundamental; (3) cantos/minuto: repetición de los cantos en el lapso de un minuto; (4) Notas canto: número de unidades acústicas de un determinado patrón de amplitud reconocido en los cantos; (5) Duración de los cantos: tiempo desde el inicio hasta el final de un canto, medido con el analizador de forma de onda, (6) Intervalos entre cantos: tiempo transcurrido entre canto y canto. Las definiciones y mediciones realizadas en las variables acústicas se basaron en los términos propuestos por Duellman y Pyles [26], Cocroft y Ryan [27]; Angulo et al. [28]; Díaz y Cádiz [29] y Batallas y Brito [24].
RESULTADOS
Pristimantis caniari sp. nov.
Fig. 1-4
Holotipo: DHMECN 10341, macho adulto, colectado en Chacapata, cerca de la Laguna de Culebrillas, Parque Nacional Sangay, (2.404824°S, 78.833850°W, 3920 m; Datum WGS84; Fig. 5), parroquia Juncal, cantón Cañar, provincia de Cañar, República del Ecuador, el 27 de junio del 2009, por Salomón Ramírez Jaramillo.

Pristimantis caniari sp. nov. DHMECN 10341, macho, holotipo (LRC = 24,4 mm). (A) Coloración dorsal en vida; (B) Coloración ventral en vida. Fotografías: Salomón M. Ramírez-Jaramillo.
Paratopotipos: DHMECN 10342-10345 machos adultos con los mismos datos del holotipo.
Etimología: El epíteto específico “caniari” es utilizado como sustantivo en aposición, y hace alusión al pueblo y cultura Cañari. Los Cañaris fueron antiguos habitantes en las tierras frías de páramo en las provincias de Cañar y Azuay. El nombre rinde honor a esta cultura andina, cuyas creencias y tradiciones aún prevalecen en el tiempo.
Nombre común sugerido: Cutín Cañari
Nombre común en inglés sugerdio: Cañari Robber Frog
Asignación genérica: Asignamos a la nueva especie dentro del género Pristimantis por presentar cabeza tan ancha como el cuerpo; membrana timpánica diferenciada, crestas craneales ausente; procesos dentígeros del vómer presentes; condición “S” del músculo aductor; discos terminales en los dígitos expandidos; con surcos circunmarginales bien definidos y falanges terminales en forma de “T”; dedo I de la mano más corto que el dedo y II; de V más largo que el dedo III; tubérculos subarticulares no proyectados hacia afuera; textura de la piel del dorso variable; vientre aerolado [2].
Diagnosis: Pristimantis caniari se distingue de sus congéneres por la siguiente combinación de caracteres: (1) textura de la piel dorso y flancos finamente granular con distintivas verrugas aplanadas, con una hilera de verrugas aplanadas dispuestas paravertebralmente, pliegues dorsolaterales ausentes, piel del vientre fuertemente areolada; piel de la región internasal lisa; pliegue discoidal débilmente definido; (2) membrana y anillo timpánico presente, equivalente al el 56% del diámetro del ojo, redondo; en contacto con un pliegue supratimpánico prominente; (3) hocico corto, redondeado en vista dorsal y de perfil; (4) párpado superior con uno a dos tubérculos redondeados pequeños, más estrecho que la distancia interorbital, cresta craneal ausente; (5) procesos dentígeros del vómer de contorno triangular, con 4 a 7 dientes; (6) machos con hendiduras bucales sublinguales, saco gular y almohadillas nupciales presentes; (7) dedo I de la mano más corto que el dedo II; discos ligeramente más anchos que los dígitos, levemente truncados de los dedos II al IV; (8) dedos de la mano con rebordes cutáneos laterales presentes; (9) tubérculos ulnares presentes, aplanados, dispuestos en una hilera longitudinal a manera de pliegue; (10) talón con varios tubérculos redondeados, borde externo del tarso con verrugas aplanadas; pliegue tarsal interno presente, se extiende a las 3⁄4 partes del tarso; (11) tubérculo metatarsal interno ovalado, 2-3 veces el tamaño del tubérculo metatarsal externo, redondeado; tubérculos supernumerarios presentes; (12) dedos del pie con rebordes cutáneos laterales; sin membranas interdigitales; dedo V más largo que el dedo III y no se extiende más allá del tubérculo subarticular distal del dedo IV; discos del mismo tamaño que los discos de la mano; (13) coloración dorsal desde café claro, café oscuro a gris oscuro, con manchas café oscuras, negras o blancas; flancos homogéneamente café con verrugas blancas a crema (blancas en vida); garganta y vientre en fondo café claro hasta café oscuro con distintivas verrugas pigmentadas de blanco; línea labial completa o interrumpida de color crema; superficies ocultas de las ingles con coloración café claro (rojizo en vida), iris dorado con finas reticulaciones negras; (14) longitud rostro-cloacal en machos 24,1-24,9 mm (!”=24,5, n= 5), hembras desconocidas.
Comparación con especies similares: Las principales diferencias con los miembros del grupo de P. orcesi (sensu Hedges et al. [2]) se resumen en el Tabla 3. En su morfología externa Pristimantis caniari es reminiscente a las especies del grupo de P. orcesi. Dentro del grupo de especies de P. orcesi, P. obmutescens, P. racemus, P. simoteriscus y P. simoterus (Fig. 6), distribuidas en la Cordillera Central de Colombia, pueden resultar similares a P. caniari (caracteres entre paréntesis) por la presencia de verrugas aplanadas en el dorso y por exhibir un vientre fuertemente areolado. Sin embargo, las cuatro especies muestran procesos dentígeros del vómer de contorno oblicuo (triangular) y coloración del vientre crema con reticulaciones café-rojizas a gris uniforme en P. obmutescens, blanco con puntos y reticulaciones cafés en P. racemus, café rojizo pálido en P. simoterus y café pálido en P. simoteriscus (café claro a café oscuro con distintivas verrugas pigmentadas de blanco); P. obmutescens presenta tímpano oculto detrás de la piel, ausencia de hendiduras bucales y saco gular, ausencia de almohadillas nupciales (todos los caracteres presentes); las hendiduras bucales, saco gular y almohadillas nupciales están ausentes en P. racemus (presentes); P. simoterus posee débiles crestas craneales (ausentes), ausencia de almohadillas nupciales (presentes), tubérculos ulnares ausentes (aplanados, dispuestos en una hilera longitudinal a manera de pliegue); P. simoteriscus presenta el párpado superior sin tubérculos (uno a dos redondeados, pequeños), ausencia de hendiduras bucales y almohadillas nupciales (presentes), tubérculos ulnares ausentes o pobremente definidos (aplanados, dispuestos en una hilera longitudinal a manera de pliegue).

(A) vista dorsal. (B) vista ventral. (C) vista de perfil de Pristimantis caniari sp. nov., DHMECN 10341, macho, holotipo (LRC = 24,4 mm).
Las especies ecuatorianas del grupo de Pristimantis orcesi: P. orcesi, P. thymelensis, y P. ortizi (Fig. 7) pueden ser semejantes a P. caniari (caracteres entre paréntesis). No obstante, P. orcesi se distingue por carecer de procesos dentígeros del vómer (presentes), tubérculos subarticulares redondeados, visibles y pronunciados (ligeramente elevados); discos digitales más anchos que los dígitos (ligeramente más anchos); tubérculos ulnares ausentes (presentes); tubérculo metatarsal interno 3-5 veces el tamaño del externo (2-3 veces); coloración dorsal en vida de café a negro con o sin verrugas blancas en los flancos, vientre café claro (dorso desde café oscuro, café con manchas blancas, hasta café chocolate; flancos homogéneamente café oscuros con verrugas blancas; garganta y vientre en fondo café claro hasta café oscuro con distintivas verrugas pigmentadas de blanco).
Pristimantis thymelensis se diferencia de P.caniari (caracteres entre paréntesis) por presentar el tímpano oculto bajo la piel (visible externamente); tubérculos en el párpado superior ausentes (uno a dos tubérculos pequeños redondeados); procesos dentígeros del vómer de contorno oblicuo (triangular); tubérculos ulnares prominentes (aplanados dispuestos en hilera longitudinal a manera de pliegue); tubérculo tarsal interno presente (pliegue tarsal interno); tarso con tubérculos redondeados (con verrugas aplanadas); tubérculo metatarsal interno 5-6 veces el tamaño del externo (2-3 veces); vientre gris cremoso a amarillo marrón y densamente reticulado con café o negro (café claro a café oscuro con distintivas verrugas pigmentadas de blanco). Por otro lado, P. ortizi se diferencia de la nueva especie por presentar la piel del dorso areolada (fuertemente areolada), procesos dentígeros del vómer ausentes o reducidos (presentes, triangulares), almohadillas nupciales ausentes (presentes), tubérculos ulnares usualmente ausentes, si presentes pequeños (aplanados, dispuestos en una hilera longitudinal a manera de pliegue); vientre blanco con motas café oscuras (café claro a café oscuro con distintivas verrugas pigmentadas de blanco).
Otra especie de los Andes centro-sur del Ecuador que comparte el carácter de verrugas aplanadas en el dorso y flancos es P. riveti (Fig. 7), sin embargo, se distingue de P. caniari por presentar pliegues dorsolaterales evidentes (ausentes); procesos dentígeros del vómer redondos (triangulares), tubérculos ulnares presentes, poco definidos (presentes, aplanados, dispuestos en una hilera longitudinal a manera de pliegue), tubérculo metatarsal externo cónico (redondeado), pliegue tarsal interno ausente (presente); saco gular en machos amarillo y vientre de blanco perla a rosa pálido con reticulaciones café (garganta y vientre en fondo café claro hasta café oscuro con distintivas verrugas pigmentadas de blanco).

Pristimantis caniari sp. nov. DHMECN 10341, macho, holotipo. (A) Detalle de las extremidades; (B) Detalle de la cabeza en vista de dorsal y de perfil.
Descripción del holotipo: Macho adulto, cabeza más larga que ancha; hocico corto, redondeado en vista dorsal y de perfil (Fig. 3); distancia ojo-narina 13% de la LRC; canto rostral ligeramente cóncavo, narinas débilmente dirigidas lateralmente (Fig. 3); área interorbital ligeramente cóncava, más ancha que el párpado superior, equivalente al 62% de la distancia interorbital; cresta craneal ausente; párpado superior con uno a dos tubérculos redondeados pequeños (Fig. 3), párpado más angosto que el espacio interorbital; membrana timpánica diferenciada de la piel que la rodea, anillo timpánico evidente, redondo, margen superior en contacto con un pliegue supratimpánico grueso y verrugas aplanadas, tímpano visible parcialmente en visita dorsal, diámetro del tímpano 63% del diámetro del ojo, verrugas postrictales redondeadas presentes; coanas pequeñas, ovoides de contorno, no cubiertas por el piso palatal del maxilar; procesos dentígeros del vómer presentes, triangular de contorno con 5-6 dientes, ubicados posteriomedialmente en relación a las coanas; lengua más larga que ancha, ovalada, 40% adherida al piso palatal de la boca.
Textura de la piel en la región internasal lisa, dorso y flancos finamente granular con distintivas verrugas aplanadas, con una hilera de verrugas aplanadas dispuestas paravertebralmente, pliegues dorsolaterales ausentes, piel del vientre fuertemente areolada. Pliegue discoidal débilmente definido, cloaca rodeada por algunas verrugas redondeadas y aplanadas. Brazos esbeltos, tubérculos ulnares presentes, aplanados, dispuestos en una hilera longitudinal a manera de pliegue (Fig. 3); dedos con rebordes cutáneos laterales, tubérculo palmar bifurcado, semejante a un corazón, tubérculo tenar ovalado (Fig. 3); tubérculos subarticulares redondos prominentes, tubérculos supernumerarios presentes, grandes y aplanados; almohadillas nupciales presentes, bien definidas, evidentes en el dedo I, terminaciones digitales ligeramente más anchas que los dígitos, discos definidos por surcos circunmarginales (Fig. 3).
Extremidades posteriores esbeltas; longitud de la tibia 41% de la longitud rostro- cloacal, talón con varios tubérculos redondeados, borde externo del tarso con verrugas aplanadas (Fig. 3); pliegue tarsal interno presente, se extiende a las 3⁄4 partes del tarso (Fig. 3); dedos del pie con rebordes cutáneos laterales, pronunciados basalmente en todos los dedos; tubérculos subarticulares redondos, prominentes; tubérculo metatarsal interno ovalado, 3 veces del tubérculo metatarsal externo que es redondeado; tubérculos supernumerarios presentes, grandes y aplanados (Fig. 3); discos ligeramente más anchos que los dígitos; dedo V mayor al III, no se extiende más allá del tubérculo subarticular distal del dedo IV (Fig. 3).
Coloración en preservado del holotipo: Dorso uniformemente gris azulado; flancos e ingle café con verrugas aplanadas crema; garganta y vientre café, vientre con distintivas verrugas pigmentadas de blanco; débil línea supralabial crema; dorsalmente, extremidades anteriores de color gris azulado, bordes internos de la ulna y I a III dedo café; superficies posteriores y ocultas de las extremidades posteriores café (Fig. 2).
Coloración en vida: Dorsalmente café claro y oscuro a gris oscuro, con manchas café oscuras, negras o blancas; hocico siena brillante a café oscuro, franja supralabial crema con o sin interespacios oscuros; flancos en fondo café claro a oscuro con distintivas verrugas blancas; extremidades café claro y oscuro a gris oscuro con manchas irregulares café oscuras, negras o blancas, dígitos delineados de crema a siena brillante; vientre fondo café claro a oscuro, con distintivas verrugas blancas; garganta café claro a oscuro; ingle y superficies anteriores y posteriores de los muslos café rojizo; iris dorado con finas reticulaciones negras (Fig. 1).
Medidas del holotipo (mm): longitud rostro-cloacal = 24,4; distancia órbita-narina = 3,1; longitud cefálica = 10,3; ancho cefálico = 9,7; distancia interorbital = 3,5; distancia internarinal =2,3; longitud de la tibia = 10,1; longitud del pie = 10,3; longitud de la mano = 6,9; diámetro horizontal del tímpano = 1,6; longitud horizontal del ojo = 2, 6; ancho del párpado = 2,2.
Variación: Variaciones morfométricas de la serie tipo son presentadas en la Tabla 1. El espécimen DHMECN 10341 exhibe un patrón de coloración dorsal café, el espécimen DHMECN 10344 café claro con marcas longitudinales café oscuro, el paratopotipo DHMECN 10342 presenta una coloración dorsal gris azulado con distintivas marcas blancas, mientras que el ejemplar DHMECN 10345 muestra el dorso gris azulado con marcas gris claro en “ʌ”, el holotipo por su lado presenta un patrón uniforme gris azulado. Ventralmente los paratopotipos DHMECN 10341-10342, 10344 muestran distintivas verrugas pigmentadas de crema en fondo café claro, mientras que el holotipo y el paratopotipo DHMECN 10345 exhiben dichas verrugas en fondo café oscuro (Fig. 4).

Variación dorsal y ventral en la serie tipo de Pristimantis caniari sp. nov. Primera y segunda fila de izquierda a derecha: DHMECN 10341, macho, holotipo (LRC = 24,4 mm); DHMECN 10342, macho, paratopotipo (LRC = 25,4 mm); DHMECN 10343, macho paratopotipo (LRC = 24,7 mm); DHMECN 10344, macho, paratopotipo (LRC = 25,2 mm); DHMECN 10345 macho, paratopotipo (LRC = 25,1 mm).
| Machos n=5 | |
| LRC | 24,1-24,9 (24,5±0,3) |
| ON | 2,4- 3,1 (2,8± 0,3) |
| LC | 8,7-10,3 (9,5± 0,6) |
| AC | 9,7-9,2 (9,5±0,2) |
| DIO | 3,2-3,7 (3,5±0,2) |
| DIN | 2,2-2,7 (2,4±0,2) |
| LT | 9,3-10,5 (10,0±0,5) |
| LP | 9,8-11,1 (10,7±0,7) |
| LM | 6,3- 7,8 (6,9±0,6) |
| DT | 1,4-1,7 (1,6±0,1 ) |
| DO | 2,6-3,1 (2,8±0,2 ) |
| AP | 2,1-2,2 (2,5±0,04) |
Medidas morfométricas de la serie tipo de Pristimantis caniari sp. nov. Rango de medidas (media ± desviación estándar). DON = distancia órbita-narina; LC = longitud cefálica; AC= ancho cefálico; DIO= distancia interorbital; DIN= distancia internarinal; LT= longitud de la tibia; LP= longitud del pie; LM= longitud de la mano; DT= diámetro horizontal del tímpano; DO=; AP= ancho del párpado
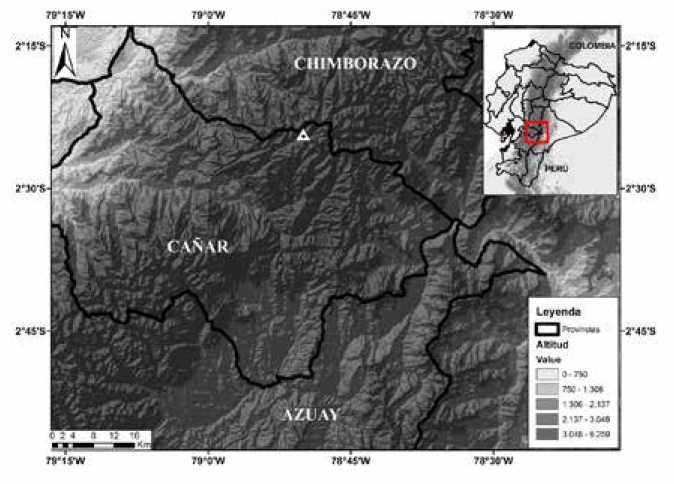
Mapa de la localidad tipo de Pristimantis caniari sp. nov., Chacapata (triángulo), Provincia de Cañar.

Especies colombianas del grupo de Pristimantis orcesi sensu Hedges et al. [2] similares a P. caniari sp. nov. (A) Pristimantis obmutescens, ICN 2087, hembra, holotipo (LRC = 31.9 mm); (B) Pristimantis racemus, UVC 8193, hembra (LRC = 33.6 mm); (C) Prsitimantis simoteriscus, ICN 22835, hembra, holotipo (LRC = 30.1); (D) Pristimantis simoterus, ICN 759, hembra, holotipo (LRC = 37.1 mm).

Especies de Pristimantis similares a P. caniari sp. nov. (A) Pristimantis orcesi, DHMECN 2903, hembra (LRC = 37,6 mm); (B) Pristimantis thymelensis DHMECN1875, hembra (LRC = 29,19 mm); (C) Pristimantis riveti, DHMECN 3084 macho (LRC = 38,5 mm).
Bioacústica: Pristimantis caniari posee cantos compuestos de una sola nota, de frecuencia modulada, cuya frecuencia dominante es de = 2,13 ± 0,02 kHz; presenta dos armónicos parciales, de los cuales el primero tiene una frecuencia de X= 4.2 ± 0.06 kHz y el segundo una frecuencia de = 6,1 ± 0,02 kHz (Fig. 8). Los cantos tienen una duración de = 207 ± 9,4 ms, con intervalos de = 11057,6 ± 3818,6 ms, emitiendo = 5,8 ± 1,5 cantos/minuto (Tabla. 2). Sus cantos son silbidos cortos, fuertes y medianamente agudos, propios de ranas terrestres Pristimantis de alturas, las vocalizaciones fueron emitidas desde pajonales en el páramo. Los cantos se escucharon a partir de las 18h20, incrementando su actividad a partir de las 19h00 hasta las 22h00, se escucharon vocalizaciones dispuestas en varios coros.
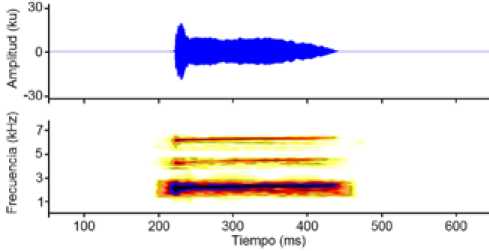
Oscilograma y espectrograma del canto de Pristimantis caniari sp. nov. Ejemplar DHMECN 10341.
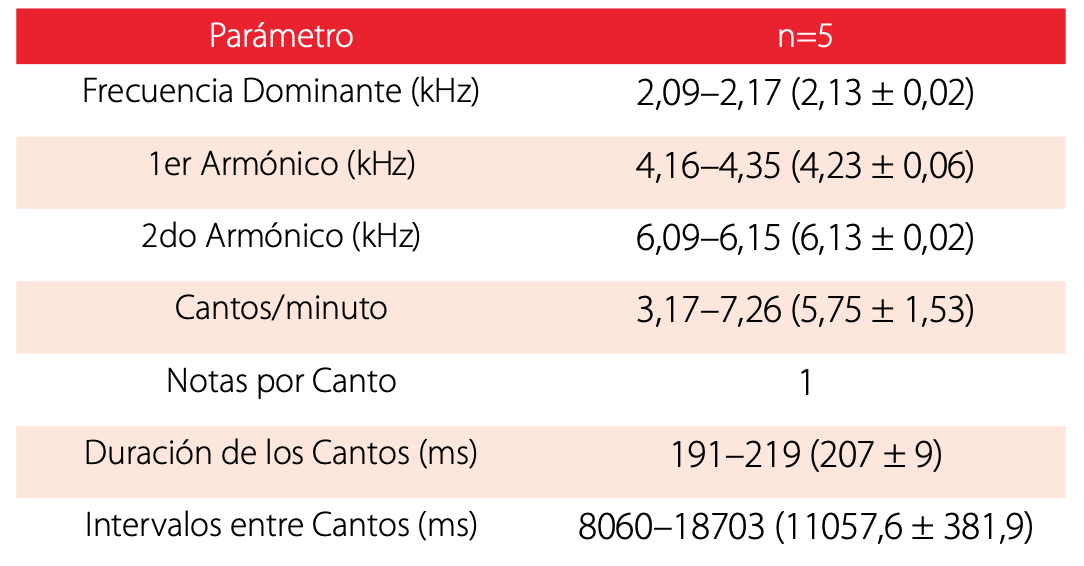
Resumen de los análisis acústicos de los cantos de Pristimantis caniari sp. nov. macho, holotipo DH- MECN 10341. De los valores calculados se detalla mínima, máxima, media y desviación estándar. Las abreviaciones utilizadas corresponden a: n= número de cantos analizados, kHz= Kilohertzios, ms= milisegundos.
Distribución e historia natural:Pristimantis caniari es conocido únicamente de su localidad tipo, en el ecosistema Herbazal de Páramo, piso bioclimático montano alto y montano alto superior [30], en la provincia de Cañar a 3920 m de elevación. La localidad se ubica en los páramos del piso zoogeográfico Altoandino [31]. Los especímenes fueron colectados en vegetación herbácea, dentro de pajonales (Calamagrostis sp.), entre 10-30 cm del suelo, se capturaron 5 individuos vocalizando durante la noche. Tres muestras fueron tomadas mediante frotis para detectar la presencia Batrachochytrium dendrobatidis según la técnica de Boyle [32], mostrando resultados negativos para el patógeno. Las principales amenazas observadas durante los días de muestreo fueron: la quema de páramo en ciertas épocas del año y el sobre pastoreo en la zona. Se identifican como sus potenciales depredadores a las aves rapaces Falco sp., Phalcoboenus carunculatus, Geranoetus melanogaster (S. Ramírez-Jaramillo. com. pers. 2016).
DISCUSIÓN
Históricamente los especialistas de las ranas terrestres Pristimantis han usado los grupos o asambleas de especies como una propuesta preliminar y putativa para entender el componente filogenético del género [2, 33-35]. Sin embargo, la incertidumbre sobre las relaciones filogenéticas se mantiene para la mayoría de especies [23], debido a que la búsqueda de una congruencia de caracteres morfológicos diagnósticos, entre los grupos putativos y los resultados genéticos obtenidos en las filogenias propuestas por Hedges et al. [2], Pyron y Wiens [36], y Padial et al. [23] es todavía inconclusa. El uso de estas hipótesis puede proveer evidencias primarias que faciliten la confrontación de linajes morfológicamente similares. Consideramos que Pristimantis caniari es reminiscente en su morfología externa a las especies de ranas terrestres del grupo de P. orcesi (sensu Hedges et al. [2]), por la presencia de las siguientes características: cuerpo robusto, cabeza estrecha, hocico corto, extremidades posteriores moderadamente pequeñas; I dedo de la mano más corto que el dedo II; dedo V del pie mucho más largo que el dedo III, extendiéndose al borde proximal del tubérculo subarticular del dedo IV; discos digitales expandidos; membrana y anillo timpánico diferenciados (ausente en P. thymelensis); cresta craneal ausente; hendiduras bucales presentes; procesos vomerinos odontóforos presentes (ausentes en P. orcesi) [2].
La validez del grupo de especies de Pristimantis orcesi es todavía incierto. Originalmente Lynch [34] reconoció al grupo como una asamblea de especies del grupo de Pristimantis unistrigatus asociando cinco especies (P. obmutecens, P. orcesi, P. racemus, P. simoterus y P. thymelensis) de la cordillera central de Colombia y los Andes norte de Ecuador. Posteriormente Lynch y Duellman [1] edificarían el grupo de especies de P. orcesi, asociándolo a la serie de especies de P. martinicensis, y más tarde Guayasamín [37], hipotetizó la monofília del grupo con la sinapomorfía de reducción de fontanelas frontoparietales. Por otro lado, Hedges et al. [2] define el grupo y lo mantiene a pesar de que la evidencia filogenética con base molecular ha identificado la parafilia del grupo con muestras de tan solo dos de las ocho especies asignadas al grupo [2], y Padial et al. [23] asigna a todos los miembros del grupo de P. orcesi (sensu Hedges et al. [2]) dentro del género Pritimantis sin grupo de especies, basándose en la presencia de P. thymelensis junto a representantes del también parafilético grupo de P. myersi.
Por lo tanto, la ausencia de evidencias moleculares nos impide proponer con certeza que P. caniari presente relaciones de ancestro-descendencia con los congéneres del grupo de P. orcesi, ya que podría estar asociado a dos clados parafiléticos. No obstante, comparamos a la nueva especie con los miembros del grupo de P. orcesi (sensu Hedges et al. [2]), al ser el grupo de especies morfológicamente similar a P. caniari.
El canto de P. caniari presenta un silbido lánguido y melodioso cuyo patrón es comparable con los cantos de ciertas especies de las zonas alto andinas (e.g. grupo de especies P. myersi y P. phoxocephalus), diferenciando su sonoridad con los característicos “tics” metálicos de otros Pristimantis (sonoridad onomatopéyica muy común en zonas altas) (Batallas. com. pers. 2016). No existe una descripción formal del canto de especies afines a P. caniari, por lo tanto, desconocemos patrones acústicos de las especies asociadas a la nueva especie. La bioacústica en Ecuador se ha venido desarrollando de manera sigilosa, limitando sus estudios a complementar las descripciones de especies nuevas [12]. Es necesario acrecentar esfuerzos en el componente acústico de anuros, para de ese modo trabajar complementariamente con herramientas útiles, que permitan profundizar el estudio de relaciones filogenéticas de grupos conflictivos.
Agradecimientos
Dejamos constancia de nuestro agradecimiento al Ministerio de Ambiente de Ecuador por facilitar el permiso de investigación N°001-09-IC-FAU-FLO-DNB-/MA para la colecta de especímenes. La revisión del material tipo de Pristimantis en Colombia, contó el apoyo económico del Jardín Botánico Basel a través de Heinz Schneider y de Lou Jost, Javier Robayo y Juan P. Reyes-Puig de Fundación Ecomiga. John D. Lynch y Raúl Sedano brindaron las mejores condiciones durante la visita de MYM a las colecciones del Instituto de Ciencias Naturales (ICN) de la Universidad de Colombia y la Colección Herpetológica de la Universidad del Valle del Cauca (UVC). El trabajo de los autores se enmarca en el Programa de Investigación Red Terrarana del Ecuador, respaldado por el INABIO.
Referencias
Lynch, J. D., & Duellman, W. E. (1997). Frogs of the genus Eleutherodactylus (Leptodactylidae) in western Ecuador: Systematics, Ecology, and Biogeography. University of Kansas, Museum of Natural History, Special Publication, 23, 1–236. doi: https://doi.org/10.5962/bhl.title.7951
Hedges, S., Duellman, W., & Heinicke, W. E. (2008). New World direct-developing frogs (Anura: Terrarana): Molecular phylogeny, classification, biogeography, and conservation. Zootaxa 1737,1-182.
Frost, D. R. (2016, Diciembre 5). Amphibian Species of the World: an Online Reference. Versión 6.0. American Museum of Natural History. URL: http://research.amnh.org/herpetology/amphibia/index.html
Ron, S. R., Guayasamín, J. M., Yánez-Muñoz, M. H., Merino-Viteri, A. Ortiz, D. A., & Nicolalde, D. A. (2016, Diciembre 16). AmphibiaWebEcuador. Version 2016.0. Museo de Zoología, Pontificia Universidad Católica del Ecuador. URL: http://zoologia.puce.edu.ec/Vertebrados/anfibios (Error 5: El enlace externo http://zoologia.puce.edu.ec/Vertebrados/anfibios. debe ser una URL) (Error 6: La URL http://zoologia.puce.edu.ec/Vertebrados/anfibios. no esta bien escrita)
Yánez-Muñoz, M. H., Sánchez-Nivicela, J.C. & Reyes-Puig, C. 2016. Tres nuevas especies de ranas terrestres Pris- timantis (Anura: Craugastoridae) de la provincia de El Oro, Ecuador. Avances en Ciencias e Ingenierías, (8) 1, 5-25. DOI: http://dx.doi.org/10.18272/aci.v8i1.455
Brito, J., Ojala Barbour, R., Batallas D. & Almedáriz, A. (2016). A new species of Pristimantis (Amphibia: Strabo- mantidae) from the cloud forest of Sangay National Park, Ecuador. Journal of Herpetology, 50 (2), 337-344. doi: http://dx.doi.org/10.1670/13-103
Navarrete, M. J., Venegas, P. & Ron, S. R. (2016). Two new species of frogs of the genus Pristimantis from Llanganates National Park in Ecuador with comments on the regional diversity of Ecuadorian Pristimantis (Anura, Craugastoridae). ZooKeys, 593, 139-162. doi: https://doi.org/10.3897/zookeys.593.8063
Reyes-Puig, J. P., Reyes-Puig, C., Pérez L., M. & Yánez-Muñoz, M. H. (2015). Dos nuevas especies de ranas Pristimantis (Craugastoridae) de la cordillera de los Sacha Llanganatis, vertiente oriental de los Andes de Ecuador. Avances en Ciencias e Ingenierías, (7) 2, B61-B74. doi: http://doi.org/10.18272/aci.v7i2.258
Yánez-Muñoz, M. H., Reyes-Puig, C., Bejarano-Muñoz, P. & Ron, S. R. (2015). Otra nueva especie de Pristimantis (Anura: Terrarana) de las estribaciones occidentales del Volcán Pichincha, Ecuador. Avances en Ciencias e Ingenierías, (7) 2, B52- B60. doi: http://doi.org/10.18272/aci.v7i2.257
Arteaga, A. F., Pyron, R. A., Peñafiel, N., Romero-Barreto, P., Culebras, J., Bustamante, L. M., Yánez-Muñoz, M. H. & Guayasamín, J. M. (2016). Comparative phylogeography reveals cryptic diversity and repeated patterns of cladogenesis for amphibians and reptiles in northwestern Ecuador. PLoS ONE, 11 (4). DOI: http://dx.doi.org/e0151746. doi: http://doi.org/10.1371/journal.pone.0151746
Guayasamin, J. M., Krynak, T., Krynak, K., Culebras, J. & Hutter, C. R. (2015). Phenotypic plasticity raises questions for taxonomically important traits: a remarkable new Andean rainfrog (Pristimantis) with the ability to change skin texture. Zoological Journal of the Linnean Society, 173, 913-928. doi: http://doi.org/10.1111/zoj.12222
Brito, J., Batallas, D. & Velalcazar, D. (2014). Nueva especie de rana terrestre del género Pristimantis (Anura: Craugastoridae), meseta de la Cordillera del Cóndor. Papeis Avulsos de Zoologia, 54:435-446. doi: http://doi.org/10.1590/0031-1049.2014.54.30
Reyes-Puig, J. P., Reyes-Puig, C., Ramirez-Jaramillo, S. R., Perez, M. B. & Yánez-Muñoz, M. H. (2014). Tres nuevas especies de ranas terrestres Pristimantis (Anura: Craugastoridae) de la cuenca alta del Río Pastaza, Ecuador. Avances en Ciencias e Ingenierías, 6 (2), B51-B62. doi: http://doi.org/10.18272/aci.v6i2.179
Lynch, J. D. y Duellman, W. E. (1980). The Eleutherodactylus of the Amazonian slopes of the Ecuadorian Andes (Anura: Leptodactylidae). The University of Kansas, Museum of Natural History, Miscellaneous Publications 69:1-86. doi: https://doi.org/10.5962/bhl.title.16222
Lehr, E. & Coloma, L. (2008). A minute new Ecuadorian Andean Frog (Anura: Strabomantidae, Pristimantis). Herpetologica, 64(3):354-367. doi: https://doi.Org/10.1655/07-089.1
Bustamante, M. & Mendelson, J. (2008). A new frog species (Strabomantidae: Pristimantis) from the High Andes of Southeastern Ecuador. Zootaxa, 1820:49-59. doi: http://dx.doi.org/10.5281/zenodo.182997
Brito, J. & Pozo-Zamora, G. (2013). Una nueva especie de rana terrestre del género Pristimantis (Amphibia: Craugastoridae), de la Cordillera de Kutukú, Ecuador. Papeís Avulsos de Zoología, 53(24), 315-325. doi: http://doi.org/10.1590/0031-1049.2014.54.30
Guayasamin, J. M. & Arteaga, A. F. (2013). A new species of the Pristimantis orestes group (Amphibia: Strabomantidae) from the high Andes of Ecuador, Reserva Mazar. Zootaxa 3616:345-346. doi: http://doi.org/10.11646/zootaxa.3616.4.3
Arteaga-Navarro, A. F. & Guayasamin, J. M. (2011). A new frog of the genus Pristimantis (Amphibia: Strabomantidae) from the high Andes of Southeastern Ecuador, discovered using morphological and molecular data. Zootaxa 2876: 17-29.
Urgilés, V. L., Sánchez-Nivicela, J. C., Nieves, C. & Yánez-Muñoz, M. H. 2014. Ranas terrestres en los ecosistemas surandinos de Ecuador I: Dos nuevas especies de Pristimantis (Anura: Craugastoridae) de la ladera oriental. Avances en Ciencias e Ingenierías, 6 (1), B51-B59. doi: http://doi.org/10.18272/aci.v6i1.159
Duellman, W., & Lehr, E. (2009). Terrestrial breeding frogs (Strabomantidae) in Peru. Germany: NTV Science.
Heinicke, M. P., Lemmon, A. R., Lemmon, E. M., McGrath, K., & Hedges, S. B. 2018. Phylogenomic support for evolutionary relationships of New World direct-developing frogs (Anura: Terraranae). Molecular phylogenetics and evolution, 118, 145-155. doi: https://doi.org/10.1016/j.ympev.2017.09.021
Padial, J., Grant, T., & Frost, D. (2014). Molecular systematics of Terraranas (Anura: Brachycephaloidea) with an assessment of the effects of alignment and optimality criteria. Zootaxa 3825, 1-132.
Batallas, D. & Brito, J. (2014). Nueva especie de rana del género Pristimantis del grupo lacrimosus (Amphibia: Craugastoridae) del Parque Nacional Sangay, Ecuador. Papéis Avulsos de Zoologia, 54:51- 62. doi: http://doi.org/10.1590/0031-1049.2014.54.05
Charif, R.A., Waack, A.M. & Strickman, L.M. (2010). Raven Pro 1.4 User’s Manual. Cornell Lab of Ornithology, Ithaca, NY.
Duellman, W. E. & Pyles, R. A. (1983). Acoustic resource partitioning in anuran communities. Copeia, 1983:639-649. URL: http://www.jstor.org/stable/1444328
Cocroft, R.B. & Ryan, M.J. (1995). Patterns of advertisement call evolution in toads and chorus frogs. Animal Behaviour, 49:283-303. doi: https://doi.org/10.1006/anbe.1995.0043
Angulo, A. (2006). Fundamentos de bioacústica y aspectos prácticos de grabaciones y análisis de cantos. En: Angulo, A.; Rueda- Almondacid, J.V.; Rodriguez-Mahecha J.V. & E. Marca. (Eds). Técnicas de inventario y monitoreo para los anfibios de la Región Tropical Andina. Conservación Internacional, Serie Manuales de Campo, Bogotá, Colombia (pp. 93–134).
Díaz, L.M. & Cádiz, L.M. (2007). Guía descriptiva para la identificación de las llamadas de anuncio de las ranas cubanas del género Eleutherodactylus (Anura: Leptodactylidae). Herpetotropicos, 3:100-122.
Ministerio del Ambiente del Ecuador. (2012). Sistema de clasificación de los ecosistemas del Ecuador continental. Quito: Subsecretaría de Patrimonio Natural.
Albuja, L., Ibarra, M., Urgilés, J., & Barriga, R. (1980). Estudio Preliminar de los Vertebrados Ecuatorianos. Quito-Euador: Escuela Politécnica Nacional.
Boyle, D.G., Boyle, D.b., Olsen V., Morgan, J.A.T. & Hyatt, A.D. (2004) Rapid quantitative detection of chytridiomycosis (Batrachochytrium dendrobatidis) in amphibian samples using real-time Taqman PCR assay. Diseases of Aquatic Organisms, 60: 141-148. doi: http://doi.org/10.3354/dao060141
Lynch, J. D. (1976). The species groups of the South American frogs of the genus Eleutherodactylus (Leptodactylidae). Occ. Pap. Mus. Nat. Hist. Univ. Kansas, 61: 1–24. URL: http://biostor.org/reference/50169
Lynch, J. D. (1981). Leptodactylid frogs of the genus Eleutherodactylus in the Andes of Northern Ecuador and adjacent Colombia. Mis. Pub. Mus. Nat. Hist. Univ. Kansas, 72: 1–46. doi: https://doi.org/10.5962/bhl.title.16289
Lynch, J. D. D. (1980). The Eleutherodactylus of the Amazonian slopes of the ecuadorian Andes (Anura: Leptodactylidae). Mis. Pub. Mus. Nat. Hist. Univ. Kansas, 59: 75-77. doi: https://doi.org/10.5962/bhl.title.16222
Pyron, R. A., & Wiens, J. J. (2011). A large-scale phylogeny of Amphibia including over 2800 species, and a revised classification of extant frogs, salamanders, and caecilians. Molecular Phylogenetics and Evolution, 61(2), 543-583. doi: https://doi.org/10.1016/j.ympev.2011.06.012
Guayasamin, J. M. 2004. The Eleutherodactylus orcesi species group (Anura: Leptodactylidae): comparative osteology and comment on its monophyly. Herp. Monog, 18:142-174. doi: https://doi.org/10.1655/0733-1347(2004)018[0142:TEOSGA]2.0.CO;2
Apéndice
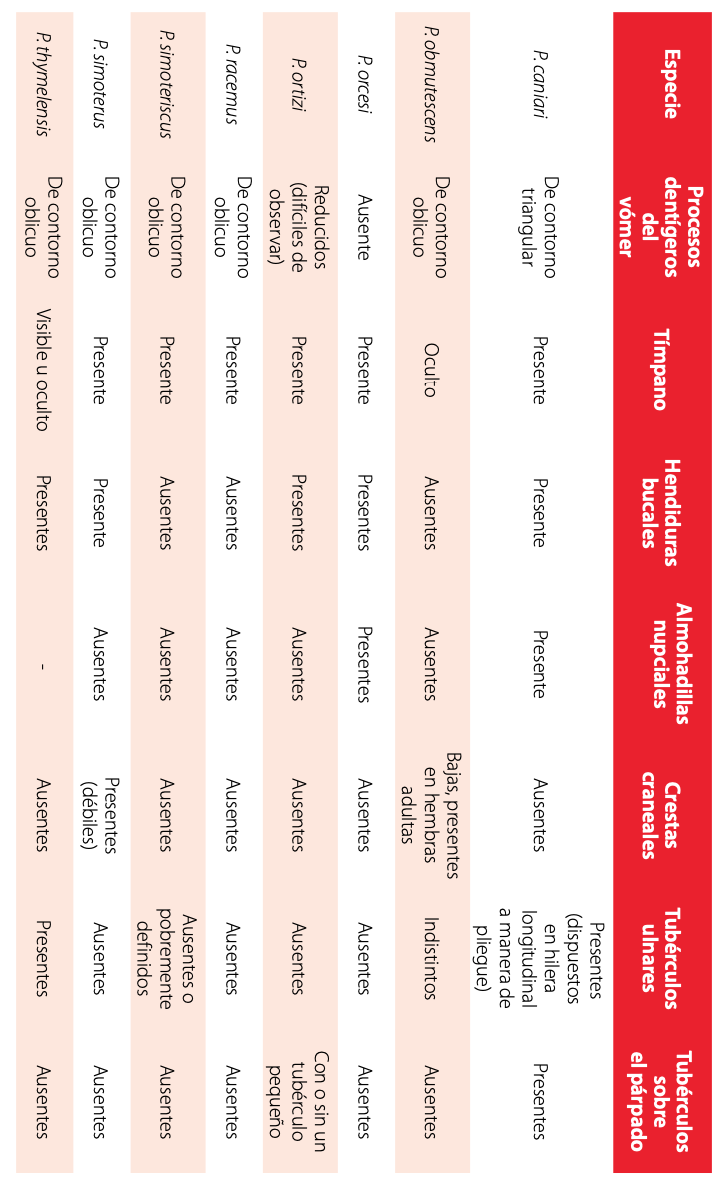
Sumario de principales caracteres utilizados para comparación dePristimantiscaniarisp. nov.
Información adicional
CONTRIBUCIONES DE LOS AUTORES
:
Salomón M. Ramírez-Jaramillo, Carolina Reyes-Puig, Diego Batallas y Mario H. Yánez-
Muñoz participaron en la colección y análisis de datos, la redacción del manuscrito y la
revisión del artículo.