

Quirópteros de una localidad piemontana amazónica del Ecuador
Chiroptera of a Foothill Amazonian locality of Ecuador
ACI Avances en Ciencias e Ingenierías
Universidad San Francisco de Quito, Ecuador
Recepción: 16 Junio 2016
Aprobación: 30 Diciembre 2017
Resumen: Se compararon los registros de murciélagos en cuatro sitios de la parroquia Simón Bolivar, Sucumbíos, Ecuador ubicados entre 640 y 1100 m de altitud en la ladera Amazónica de la Cordillera Oriental del Ecuador. Dos sitios correspondieron a bosque nativo (640 m y 1100 m) y dos fueron áreas intervenidas con cultivos y pastizal rodeadas de bosque nativo (1100 m). El muestreo se llevó a cabo en marzo de 2012 (12 noches, 400 metros de red durante 48 horas). En total, se capturaron 128 individuos distribuidos en 30 especies, de las cuales 25 pertenecen a la familia Phyllostomidae, cuatro Vespertilionidae y un Thyropteridae. Se registraron 13 especies en el bosque nativo ubicado a los 1100 m (S1), 16 en el bosque nativo ubicado en la margen derecha del río Coca a 640 m (S2), siete en pastizal rodeado de bosque nativo (S3), y 11 en el pastizal y chacra rodeados por bosque nativo (S4). La especie más abundante fue Carollia brevicauda, seguida de C. perspicillata, que representaron en conjunto el 22,7 % de la comunidad. Los sitios S2 y S4 mostraron la mayor similitud (11 %). El gremio frugívoro dominó la comunidad con más del 61 %. Se concluye que la riqueza de especies, la abundancia relativa, los índices de diversidad y la estructura trófica experimentaron una variación con relación a los sitios muestreados, probablemente debido a la combinación de bosques en buen estado de conservación con pequeñas áreas de cultivo y pasto. El bosque nativo junto al río Coca (S2) presentó los mayores valores en riqueza y abundancia. De acuerdo al estimador, el área albergaría unas 43 especies.
Palabras clave: Amazonía , andes, Chiroptera, Diversidad, Estructura trófica, Murciélagos, Sucumbíos.
Abstract: We compare records of bats at four sites along an elevational gradient (640-1100 m) on the northeastern slope of the Cordillera Real of Ecuador. Two sites were covered by native forest (640m and 1100m) and two were intervened areas with crops and pastures surrounded by native forest (1100 m). Surveys were conducted in March 2012 (12 nights, 400 meters of mistnets for 48 hours). We captured 128 individuals of 30 species in total, 25 belong to the family Phyllostomidae, four Ves- pertilionidae, and one Thyropteridae. Thirteen species were recorded at native forest at 1100 m (S1), 16 in the native forest next to the Coca River at 640 m (S2), seven in grasslands surrounded by native forest at 1100 m (S3), and 11 in grasslands and crops surrounded by native forest at 1100 m (S4). The most abundant species was Carollia brevicauda, followed by C. perspicillata, which together accounted for 22.7 % of the community. Sites S2 and S4 showed the highest similarity (11 %). Bats of the frugivorous guild dominated the community (61 %). Species richness, relative abundance, diversity indices, and trophic structure shows important variation at all sites. Native forest next to the Coca River (S2) had the highest values in richness and abundance, probably due to the combination of forest in good condition with small crops and pastures. It is estimated that the area would hold about 43 species.
Keywords: Amazonia, andes, bats, diversity, Sucumbios, trophic structure.
INTRODUCCIÓN
Los bosques de la Amazonía están considerados como altamente diversos [1]. Esta gran diversidad se debe principalmente a la alta precipitación no estacional, la cercanía a los Andes, la complejidad de los suelos, la abundancia de ríos y una particular historia biogeográfica, que crean un mosaico muy variado de hábitats [2-4]. La Amazonía, por su alta concentración de especies y amenaza, está considerada como uno de los 10 puntos calientes de biodiversidad (hotspots) en los trópicos [5]. La riqueza biológica de la Amazonía ecuatoriana es, sin duda, una de las más importantes de la región [6, 7]; más del 50 % de los mamíferos de Ecuador se encuentran en la Amazonía [8]. En Ecuador, los quirópteros están entre los grupos más diversos con 171 especies [9], lo que constituye casi el 41 % de los mamíferos registrados (414 especies).
Los murciélagos son de gran interés desde el punto de vista biológico, ecológico y de conservación. Sus funciones en los ecosistemas ha sido remarcada pues la mayoría son dispersores de semillas y polinizadores [1014] y controladores de las poblaciones de insectos [15]. El análisis e interpretación ecológica de las comunidades de murciélagos desde su función provee una nueva perspectiva y valoración de estos organismos en el ambiente [16-18]. Este trabajo se enfoca en caracterizar y comparar la comunidad de quirópteros presentes en cuatro sitios en la ladera Amazónica norte de la Cordillera Oriental del Ecuador. Como parte de esta caracterización procedemos a identificar y comparar los grupos funcionales/categorías tróficas de los quirópteros presentes en los hábitats estudiados.
MATERIALES Y MÉTODOS
Área de estudio
Los sitios de muestreo se ubican en la provincia de Sucumbíos, cantón Gonzalo Pizarro, parroquia Simón Bolívar (Fig. 1). Los sitios se establecieron entre el poblado Simón Bolívar y el río Coca, cubriendo un rango altitudinal entre 1200 m y 640 m. El ingreso se realizó por la vía hacia la casa de máquinas del Proyecto Hidroeléctrico Coca Codo Sinclair. El área incluye bosques nativos y áreas intervenidas que comprenden asociaciones de bosque con pastizal y bosque con cultivos. Los bosques de la zona alta presentan fuertes pendientes que descienden al río Coca; en algunos lugares hay pequeñas superficies planas y onduladas. Los márgenes del río Coca están formados por paredes verticales y en algunos lugares terrazas planas a onduladas con afloramientos rocosos, cubiertos de bosque nativo donde predominan Iriartea deltoidea Palmas de Pambil y Wettinia maynesis Chonta o Kili. Ecológicamente, el área corresponde a la formación vegetal de Bosque Siempreverde Piemontano [19] y, de acuerdo a Sierra, corresponde a Bosques Amazónicos de Tierras Bajas (<600 m de altitud), Bosque Siempreverde Premontano (600 m a 1300 m de altitud) y los Bosques Siempreverdes de Montaña (1300–2000 m de altitud) [20]. A continuación, una descripción detallada de los sitios de muestreo:
(51) Bosque nativo en buen estado de conservación: Este bosque se encuentra en la parte alta a 1100 m de altitud (Fig. 1). El ingreso se realiza por el km 11,5 de la vía de acceso a la casa de máquinas. Este bosque es continuo y en buen estado de conservación. La topografía es irregular y se desprenden pequeñas quebradas que desembocan en el río Coca. El sotobosque es denso, el estrato medio es definido y el dosel está formado por copas de árboles grandes, aunque hay pequeños claros dispersos con vegetación pionera herbácea y arbustiva, producto de la caída de ramas y árboles viejos. El área presenta pequeñas islas de Guadua sp. Caña guadua. Las especies arbóreas sobresalientes son representantes de las familias Moraceae, Rubiaceae, Annonaceae, Lauraceae y Fabaceae. Son comunes Cecropia spp. Guarumos, Cyathea sp. Helechos arbóreos y varias especies arbustivas del género Piper. En los árboles están presentes las epifitas (Bromeliaceae), anturios (Araceae) y orquídeas (Orchidaceae). Las redes fueron colocadas en las coordenadas 0.114425oN y 77.441922 O.
(52) Bosque nativo de la margen derecha del río Coca: Está ubicado en el km 19 de la vía de acceso a la casa de máquinas (Fig. 1). El sitio de estudio fue ubicado cerca al río Coca, la vegetación está asentada en un substrato pedregoso con una capa delgada de suelo de topografía plana. Los árboles no forman un dosel continuo, la vegetación arbórea forma pequeñas islas con sotobosque denso. La topografía está formada por quebradas, pozas pequeñas y áreas inundadas de pequeña y gran extensión que se llenan de agua cuando llueve. El dosel está formado por abundantes Wettinia maynensis Palmas de kili o Chonta e Ireartea deltoidea Pambil, otras especies que caracterizan al sitio son los árboles de Ladenbergia oblongifolia Cascarillo, Cedrela odorata Cedro, Protium fimbriatum Copal y Brosimum utile Sande, también sobresale Cecropia spp. Guarumos, Tovomita wenddeliana Pulpo, Vismia spp. Achotillo, y varias especies de Miconia spp. Colca. Esta área ha sido intervenida; durante el trabajo de campo observamos vestigios de extracción selectiva de varias especies maderables, así como algunas áreas de cultivo abandonadas en las que se está regenerando la vegetación nativa. Las redes fueron colocadas en un sitio boscoso de coordenadas 0.132783oN y 77.437111oO a 640 m de altitud.
(53) Pastizal rodeado de bosque nativo: Es un pastizal pequeño de aproximadamente 1,5 ha, con muy pocos árboles aislados y que se encuentra rodeado de un bosque nativo intervenido. El bosque presenta hasta tres estratos con algunos árboles emergentes y el sotobosque es denso. Se notan algunos signos de intervención antrópica como árboles talados y tablones aserrados. La topografía es irregular, con una pendiente pronunciadaen la parte occidental. Las redes fueron colocadas enel centro del pastizal en las coordenadas 0.093383◦N y77.429592◦O a 1100 de altitud (Fig. 1).
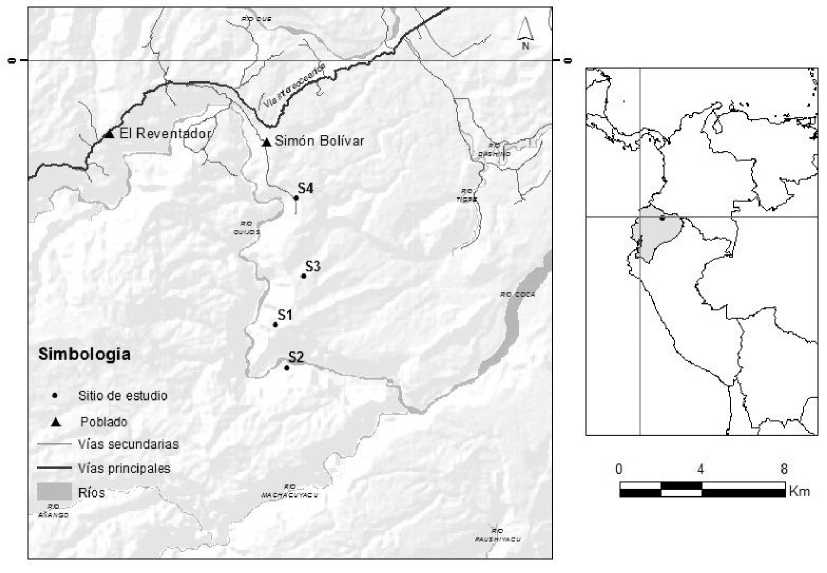
Ubicación de los sitios de muestreo de murciélagos (S) en la parroquia Simón Bolívar, cantón Gonzalo Pizarro, provincia de Sucumbíos, Ecuador.
(54) Pastizal y chacra rodeados de bosque nativo: Este sitio se encuentra al costado derecho de la vía de acceso a la casa de máquinas del Proyecto Hidroeléctrico Coca Codo Sinclair (Fig. 1). Se ingresa por el km 4, al lado derecho hay un sendero que desciende a una terraza donde los propietarios realizan actividades agropecuarias. Es un mosaico de vegetación formado por pasto con pocos árboles dispersos, cultivos, pequeñas áreas con bosque secundario y un remanente de bosque maduro en la ladera occidental de donde descienden quebradillas que desembocan en el río Coca. Las especies cultivadas en la chacra fueron Musa paradisiaca Plátano, Theobroma cacao Cacao, Coffea arabica Café, Citrus limonum Limón, Bactris gasipaes Chontaduro y Cardulovica palmata Paja toquilla. Las redes fueron colocadas entre la chacra y el pastizal, en las coordenadas 0.059542°N y 77.433011°O, a 950 m de altitud.
Métodos de campo
El trabajo de campo se realizó entre el 16 y 26 de marzo de 2013. Los murciélagos fueron capturados en redes de niebla [21] colocadas en lugares con alta probabilidad de captura. En cada sitio, se situaron ocho redes de niebla de 12 m de longitud por 2,5 m de ancho, formando dos líneas de cuatro redes cada una, ubicadas una a continuación de la otra y separadas de acuerdo a la topografía del terreno y experiencia de los investigadores. Las redes permanecieron abiertas desde las 18h15 hasta las 22h00 (tres horas 45 minutos red/noche), durante dos noches consecutivas en cada uno de los cuatro sitios de muestreo. Las redes fueron revisadas cada 20 minutos aproximadamente. Los murciélagos capturados fueron identificados en el campo usando las guías de campo de Albuja [22], Tirira [23] y el conocimiento y experiencia de los investigadores. Se tomaron medidas corporales con un calibrador digital Buffalo Tools, la masa con una Pesola, se tomaron fotografías y posteriormente los murciélagos fueron liberados.
El análisis de datos consideró la riqueza y abundancia. La riqueza es el número de especies por sitio y la suma de las especies registradas en todos los sitios estudiados. La abundancia es el número de individuos por especie. La abundancia relativa (Pi) de las especies de cada sitio y del total del muestreo se presenta en curvas de dominancia-diversidad [24]. La diversidad entre sitios se comparó con el índice Shannon [24] y Shannon Exponencial [25-26]. Se analizó la estructura funcional agrupando los registros de captura en categorías tróficas: insectívoros, frugívoros; nectarívoros y omnívoros. Se midió la similitud entre sitios mediante el índice de Jaccard [24], con esta información se realizó un análisis de conglomerados [27]. La representatividad del muestreo se presenta en una curva de acumulación de especies y el número posible de especies presentes en el área se calculó mediante un estimador (Jack-nife 1) con el programa estadístico BiodiversityPro. La categoría de amenaza de las especies se presenta de acuerdo al Libro Rojo de mamíferos del Ecuador [8] y la Lista Roja de la Unión Internacional para la Conservación de la Naturaleza (UICN) [28]. La nomenclatura taxonómica de las especies se basa en Tirira [9].

Murciélagos registrados en en la parroquia Simón Bolívar, cantón Gonzalo Pizarro, provincia de Sucumbíos, Ecuador. Las fotografías corresponden a individuos capturados en el período de trabajo de campo por los autores.
Métodos de laboratorio
Los ejemplares de difícil identificación fueron colectados para su posterior determinación en el laboratorio. Estos fueron preparados como pieles o preservados en una solución de etanol al 95 %. Estos especímenes fueron depositados en la colección de mamíferos del Museo Ecuatoriano de Ciencias Naturales del Instituto Nacional de Biodiversidad.
RESULTADOS
Riqueza, abundancia y composición total
Capturamos 128 murciélagos de 30 especies y tres familias en toda el área de estudio (Tabla 1, Fig. 2). La familia más diversa fue Phyllostomidae con 25 spp., seguida por Vespertilionidae (4 spp.) y Thyropteridae (1 spp.). La subfamilia Stenodermatinae (Phyllostomidae) fue la más diversa con 12 spp., seguida de Phyllostominae (6 spp.), Carollinae (4 spp.) y Glossophaginae (3 spp.). El 94,53 % (121 individuos) de los murciélagos capturados pertenecieron a la familia Phyllostomidae. La curva de acumulación, calculada en relación al esfuerzo de muestreo incremental con la inclusión de cada sitio adicional, se muestra en ascenso (Fig. 3). Esto sugiere que, aumentando el esfuerzo, se incrementaría el número de especies. Según el estimador (Jack-nife 1), la riqueza de especies del área de estudio llegaría a 44 especies. Se estima por lo tanto que, el número de especies registradas en este estudio representa el 69 % de los quirópteros posiblemente presentes en el área.
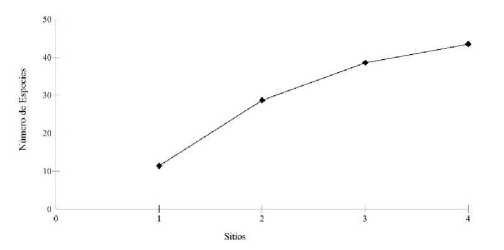
Curva de acumulación de especies de murciélagos, parroquia Simón Bolívar, cantón Gonzalo Pizarro, provincia de Sucumbíos, Ecuador, 16-26 de marzo de 2013. La curva se expresa en relación al esfuerzo de muestreo incremental con la inclusión de cada sitio adicional (S1 a S4, ver texto para descripción del esfuerzo de colección y de los sitios).
Las especies más abundantes fueron el Murciélago sedoso de cola corta Carollia brevicauda y el Murciélago común de cola corta C. perspicillata, con 17 y 12 individuos respectivamente. Estas dos especies constituyeron el 22,7 % del total de especímenes capturado. Tres especies de glosofáginos (Anoura caudifer, A. peruana y Lonchophylla robusta) acumulan una abundancia considerable (n = 19) y constituye casi el 15 % de los individuos capturados. Carollia brevicauda fue la especie dominante (Pi = 0,133) en toda el área de estudio (Fig. 4). Los murciélagos registrados en el área de estudio pertenecen a cinco gremios alimentarios: frugívoros (50 %), insectívoros (26,7 %), omnívoros (10 %), nectarívoros (10 %) y frugívoro/insectívoro (3 %) (Tabla 1, Fig. 4). Los frugívoros fueron los más abundantes con 79 individuos (61,7 %), seguidos por nectarívoros (14 %), omnívoros (11,7 %), e insectívoros (9,4 %) (Fig. 4).
De las 30 especies de murciélagos registrados, solamente dos especies están mencionadas dentro de la categoría de Casi Amenazada (NT): Sturnira oporaphilum a nivel global [28] y Sturnira magna a nivel nacional [8]. Veintitrés especies son consideradas como Preocupación menor (LC) a nivel global y nacional. Para cinco especies no se ha evaluado su estado de conservación.
Riqueza, abundancia y composición por sitio
En el Bosque nativo en buen estado de conservación (S1) capturamos 45 murciélagos de 13 especies y dos familias (Tabla 1). La familia más diversa fue Phyllostomidae con 12 spp., y solo se registró una especie de la familia Vespertilionidae. Las especies más abundantes fueron Lonchophylla robusta Murciélago anaranjado nectario y Platyrrhinus infuscus Murciélago de nariz ancha marrón, con nueve y seis individuos respectivamente. En este sitio estuvieron presentes dos especies de murciélagos Glossophaginae (Lonchophylla robusta y Anoura caudifer), la suma de sus abundancias constituye el 31 % de los murciélagos capturados. La especie dominante fue Lonchophylla robusta Murciélago nectario anaranjado (Pi=0,2) (Fig. 5). Los murciélagos registrados en S1 pertenecen a tres gremios alimentarios: frugívoros (61,5 %), insectívoros (23,1 %) y nectarívoros (23,1 %) (Tabla 1, Fig. 5). Los frugívoros fueron los más abundantes con 26 individuos (57,8 %), seguidos por los nectarívoros (31 %) y los insectívoros (11,1 %) (Fig. 5).
En el Bosque nativo de la margen derecha del río Coca (S2) capturamos 46 murciélagos de 16 especies de la familia Phyllostomidae (Tabla 1). Las especies más abundantes fueron Tonatia saurophila Murciélago grande de orejas redondas con 9 individuos, Carollia brevicauda Murciélago sedoso de orejas cortas con 5, y C. castanea Murciélago castaño de cola corta con 5. Estas tres especies constituyeron el 41 % de los murciélagos capturados y los dos primeros, el 30 %. También capturamos cuatro individuos de Anoura caudifer Murciélago longirostro con cola, que constituyen el 8 % de los murciélagos capturados. La curva de dominancia-diversidad muestra a Tonatia saurophila como la especie dominante (Pi=0,195652) (Fig. 6). Los murciélagos registrados en S2 pertenecen a cinco gremios alimentarios: frugívoros (56,3 %), omnívoros (18,8 %), insectívoros (12,5 %), nectarívoro (6,3 %) y frugívoros/insectívoros (6,3 %) (Tabla 1, Fig. 6).
| Familia / Especie | Nombre común | GA | S1 | S2 | S3 | S4 | Total |
| Phyllostomidae | |||||||
| Carollinae Carollia brevicauda | Murciélago sedoso de cola corta | Fr | 4 | 5 | 2 | 6 | 17 |
| Carollia perspicillata | Murciélago común de cola corta | Fr | 4 | 3 | 5 | 12 | |
| Carollia castanea | Murciélago castaño de cola corta | Fr | 5 | 5 | |||
| Rhinophylla pumilio | Murciélago frutero pequeño común | Fr | 1 | 4 | 5 | ||
| Glossophaginae Anoura caudifer | Murciélago longirostro con cola | Ne | 5 | 4 | 9 | ||
| Lochophylla robusta | Murciélago nectario anaranjado | Ne | 9 | 9 | |||
| Anoura peruana | Murciélago longirostro peruano | Ne | 1 | 1 | |||
| Phyllostominae Tonatia saurophila | Murciélago grande de orejas redondas | Om | 9 | 9 | |||
| Phyllostomus elongatus | Murciélago nariz de lanza menor | Om | 3 | 1 | 4 | ||
| Micronycteris minuta | Murciélago orejudo diminuto | In | 1 | 1 | 2 | ||
| Micronycteris hirsuta | Murciélago orejudo peludo | In | 1 | 1 | |||
| Gardnerycteris crenulatum | Murciélago rayado de nariz peluda | In | 2 | 2 | |||
| Phyllostomus hastatus Stenodermatinae | Murciélago nariz de lanza mayor | Om | 2 | 2 | |||
| Sturnira lilium | Murciélago pequeño de hombros amarillos | Fr | 5 | 4 | 9 | ||
| Platyrrhinus infuscus | Murciélago de nariz ancha marrón | Fr | 6 | 1 | 1 | 8 | |
| Artibeus planirostris | Murciélago frutero de rostro plano | Fr | 4 | 4 | |||
| Artibeus lituratus | Murciélago frutero grande | Fr | 1 | 1 | 1 | 3 | |
| Dermanura sp. | Murciélago frutero | Fr | 1 | 2 | 3 | ||
| Mesophylla macconnelli | Murciélago de Macconnell | Fr | 2 | 1 | 3 | ||
| Sturnira oporaphilum | Murciélago de hombros amarillos de Oriente | Fr | 3 | 3 | |||
| Uroderma bilolatum | Murciélago toldero común | Fr/In | 3 | 3 | |||
| Vampyriscus bidens | Murciélago de orejas amarillas de dos dientes | Fr | 3 | 3 | |||
| Dermanura glauca | Murciélago frutero chico plateado | Fr | 1 | 1 | 2 | ||
| Platyrrhinus nigellus | Murciélago peruano de nariz ancha | Fr | 1 | 1 | |||
| Sturnira magna | Murciélago grande de hombros amarillos | Fr | 1 | 1 | |||
| Vespertilionidae Myotis albescens | Murciélago vespertino plateado | In | 3 | 3 | |||
| Eptesicus brasiliensis | Murciélago marrón brasileño | In | 1 | 1 | |||
| Lasiurus blossevillii | Murciélago rojo del Sur | In | 1 | 1 | |||
| Myotis nigricans | Murciélago vespertino negro | In | 1 | 1 | |||
| Thyropteridae Thyroptera tricolor | Murciélago con ventosas de Spix | In | 1 | 1 | |||
| Total | 45 | 46 | 10 | 27 | 128 |
Murciélagos capturados en cuatro sitios en la parroquia Simón Bolívar, cantón Gonzalo Pizarro, provincia de Sucumbíos, Ecuador, entre el 16 y 26 de marzo de 2013. Gremio alimentario (GA): Frugívoro = Fr; Nectarívoro = Ne; Omnívoro = Om; Insectívoro = In. Sitios de estudio (S): S1 = Bosque nativo en buen estado de conservación; S2 = Bosque nativo de la margen derecha del Río Coca; S3 = Pastizal rodeado de bosque nativo; S4 = Pastizal y chacra rodeados de bosque nativo.
Los frugívoros fueron los más abundantes con 22 individuos (47,8 %), seguidos por los omnívoros con 30 % (Fig. 6).
En el Pastizal rodeado de bosque nativo (S3) capturamos 10 murciélagos de siete especies y dos familias (Tabla 1). La familia más diversa fue Phyllostomidae con 4 spp., mientras que la familia Vespertilionidae tuvo 3 spp. La especie más abundante fue Sturnira oporaphilum Murciélago de hombros amarillos de oriente con 3 individuos, y fue la especie dominante (Pi=0,3) (Fig. 7). Los murciélagos registrados en S3 pertenecen a tres gremios alimentarios: frugívoros, insectívoros y necta- rívoros, siendo los dos primeros los más representativos con 86 % de las especies de la comunidad (Tabla 1, Fig. 7). Los frugívoros fueron los más abundantes con seis individuos (60 %) (Fig. 7).
| S1 Altitud: 1.100 m | S2 Altitud: 640 m | S3 Altitud: 1.100 m | S4 Altitud: 950 m | Total 640 - 1.100 m | |
| Riqueza | 13 | 16 | 7 | 11 | 30 |
| Abundancia | 45 | 46 | 10 | 27 | 128 |
| Diversidad: Shannon | 2,328 | 2,551 | 1,834 | 2,138 | |
| Número efectivo de especies (Shannon Exponencial) | 10,26 | 12,82 | 6,26 | 8,48 | |
| No. Gremios alimentarios | 3 | 5 | 3 | 3 | 5 |
| No. Frugívoros | 26 | 22 | 6 | 25 | 79 |
| No. Nectarívoros | 14 | 4 | 1 | - | 19 |
| No. Insectívoros | 5 | 3 | 3 | 1 | 12 |
| No. Omnívoros | - | 14 | - | 1 | 15 |
| No. Frugívoro/Insectívoro | - | 3 | - | - | 3 |
| No. especies únicas | 6 | 6 | 5 | 1 | - |
Comparación de las variables analizadas entre cuatro sitios de muestreo en la parroquia Simón Bolívar, cantón Gonzalo Pizarro, provincia de Sucumbíos, Ecuador, entre el 16 y 26 de marzo de 2013.
En el Pastizal y chacra rodeados de bosque nativo (S4) capturamos 27 murciélagos de 11 especies y dos familias (Tabla 1). La familia Phyllostomidae fue la más diversa con 10 spp., mientras que la familia Tyropteridae tuvo una especie. Las especies más abundantes fueron Carollia brevicauda Murciélago sedoso de cola corta y C. perspicillata Murciélago común de cola corta, con 6 y 5 individuos respectivamente, y entre las dos constituyeron el 40,7 % de los murciélagos capturados en este sitio. La especie dominante fue Carollia brevicauda (Pi=0,22) (Fig. 8). Los murciélagos registrados en S4 pertenecen a tres gremios alimentarios: frugívoros (81,8 %), insectívoros (9,1 %) y omnívoros (9,1 %) (Tabla 1, Fig. 8). Los frugívoros fueron los más abundantes con 25 individuos (92,6 %) (Fig. 8).
La riqueza de especies entre los sitios varió de 7 a 16 especies. El Bosque de la margen derecha del río Coca (S2) tuvo la mayor riqueza con 16 especies. La abundancia de los sitios varió entre 10 y 46 individuos, con los sitios S1 y S2 mostrando las mayores abundancias de individuos. Una especie (Carollia brevicauda) fue registrada en todos los sitios, tres especies son compartidas entre tres sitios comunes y ocho especies entre dos sitios (Tablas 1-2). S2 presentó el mayor índice de diversidad (Shannon = 2,55) y el mayor número efectivo de especies (Shannon Exponencial = 12,82) (Tabla 2). S2 tuvo el mayor número de gremios, mientras que los restantes sitios presentaron tres gremios. En todos los lugares, el gremio más abundante fueron los frugívoros. Los nectarívoros fueron abundantes en S1 y los omnívoros fueron abundantes en S2 (Tablas 1-2). Cada sitio tuvo especies únicas; el mayor número (6) fue registrado en S1 y S2 (Tablas 1-2). El análisis de similitud de especies de murciélagos entre los cuatro sitios de muestreo indica que S2 y S4 son los más similares (42,11 %) (Fig. 9). Por otro lado, el análisis de similitud de relación a los grupos tróficos indica que S1 y S3 son los más similares (Fig. 9).
Notas sobre aspectos reproductivos
Entre los murciélagos capturados registramos 68 machos y 60 hembras. Observando la proporción de individuos entre machos y hembras de las especies más abundantes se observa que la proporción en Carollia brevicauda (n=17) fue 1,4:1; Carollia perspicillata (n=12) fue 1:1; Lonchophylla robusta (n=9) y Anoura caudifer (n=9) fue 1:1,3, en ambos casos, y Platyrrhynus infuscus (n=8) fue 1:1,7. Cuatro hembras se encontraban preñadas (Sturnira lilium, Carollia perspicillata, Tonatia saurophila y Phyllostomus elongatus). Además, tres individuos fueron juveniles, y corresponden a las especies Anoura caudifer, Carollia brevicauda y Platyrrhinus infuscus.
DISCUSIÓN
Las 30 especies de murciélagos registradas en la parroquia Simón Bolívar representan el 17,6 % de la riqueza del Ecuador. Si a estas especies se suman las 10 especies registradas en otros sitios dentro del Proyecto Hidroeléctrico Coca Codo Sinclair [29], constituyen 23,5 % de la riqueza de especies nacional, lo que indica una alta diversidad para este lugar. Cada sitio incrementó su riqueza con especies únicas. Esta diversidad puntual contribuye a la alta diversidad beta del área. La riqueza y la abundancia en los sitios boscosos (S1 y S2) fue mayor que en los sitios intervenidos (S3 y S4). Sin embargo, el mosaico formado por el pastizal y chacra rodeado de bosque nativo y pequeños fragmentos de bosque secundario en el interior está en tercer lugar, dejando en último lugar el pasto rodeado de bosque (S3). Este último lugar es un área pequeña y presentó la más baja diversidad taxonómica y trófica. Esto se debe a que esta área no ofrece refugio ni mayores recursos alimenticios. Los murciélagos capturados en este lugar seguramente fueron individuos que se encontraban de paso.
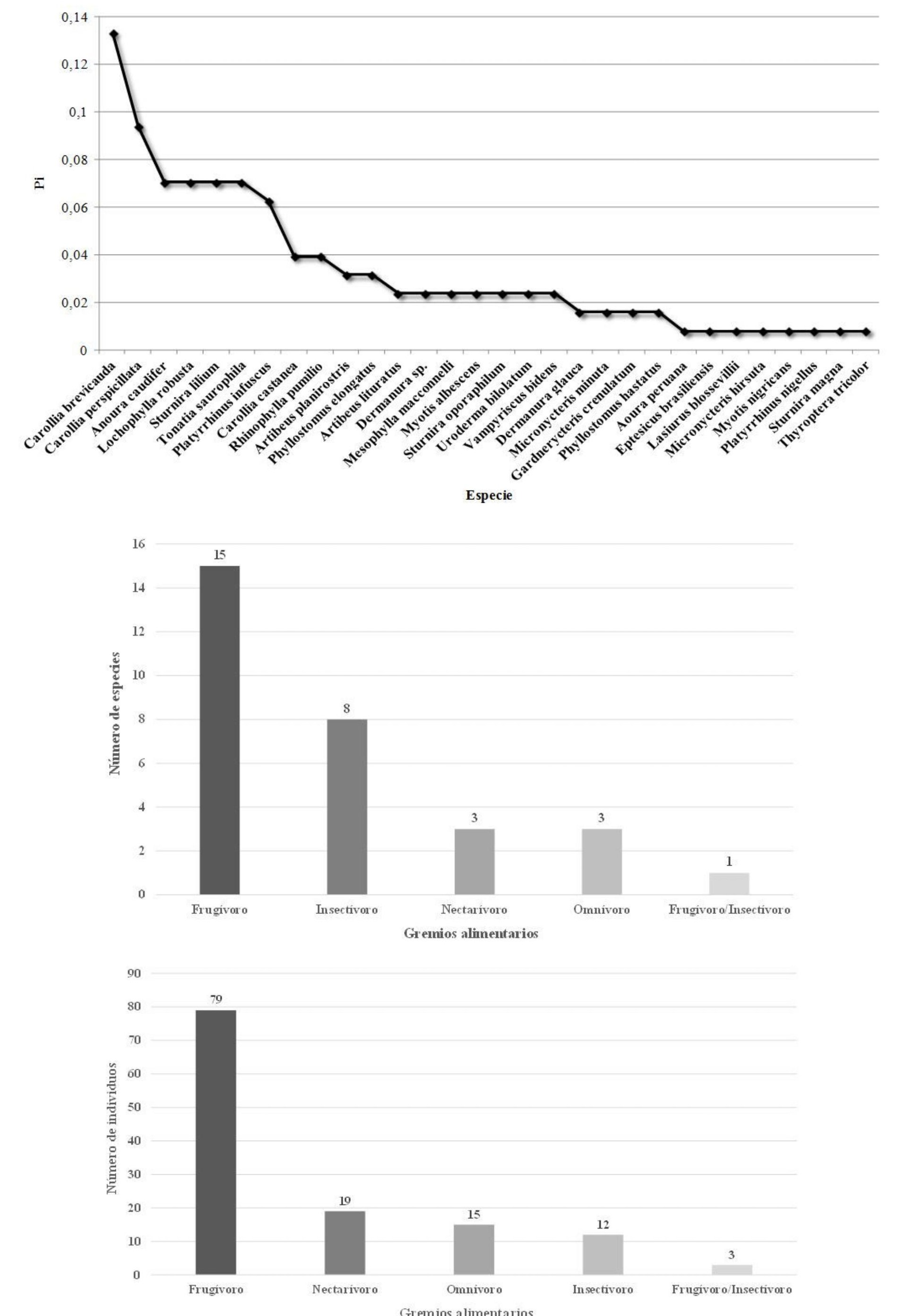
Datos de la comunidad de quirópteros en cua-tro sitios de estudio en la parroquia Simón Bolívar, cantón Gonzalo Pizarro, provincia de Sucumbíos, Ecuador, entre el 16 y 26 de marzo de 2013. Arriba: Curva de dominancia-diversidad, Pi = Proporción de individuos. Centro: Riqueza de especies. Abajo: Abundancia.

Datos de la comunidad de quirópteros capturados en Bosque nativo en buen estado de conservación (sitio S1) de la parroquia Simón Bolívar, cantón Gonzalo Pizarro, provincia de Sucumbíos, Ecuador. Arriba: Curva de dominancia-diversidad, Pi = Proporción de individuos. Centro: Riqueza de especies. Abajo: Abundancia.
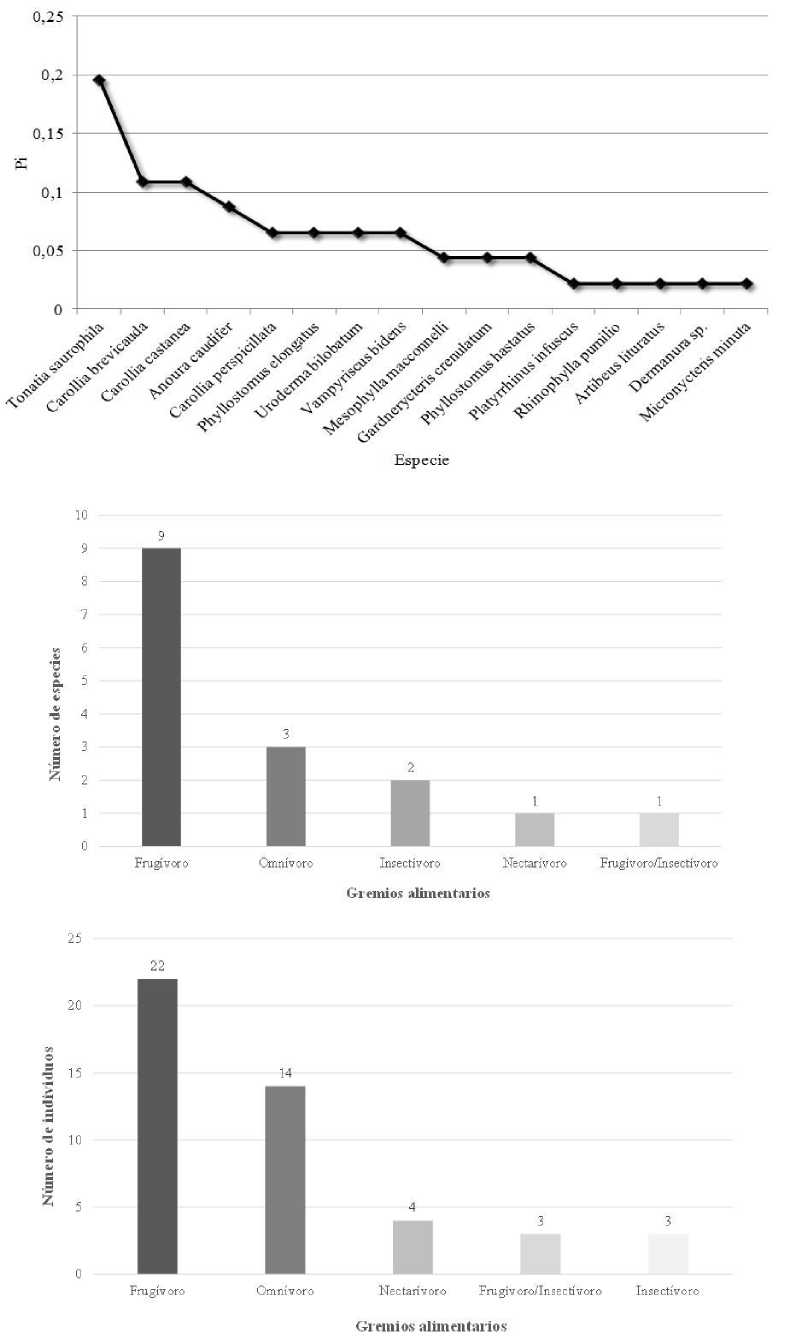
Datos de la comunidad de quirópteros capturados en Bosque nativo de la margen derecha del río Coca (sitio S2) de la parroquia Simón Bolívar, cantón Gonzalo Pizarro, provincia de Sucumbíos, Ecuador. Arriba: Curva de dominancia-diversidad, Pi = Proporción de individuos. Centro: Riqueza de especies. Abajo: Abundancia.
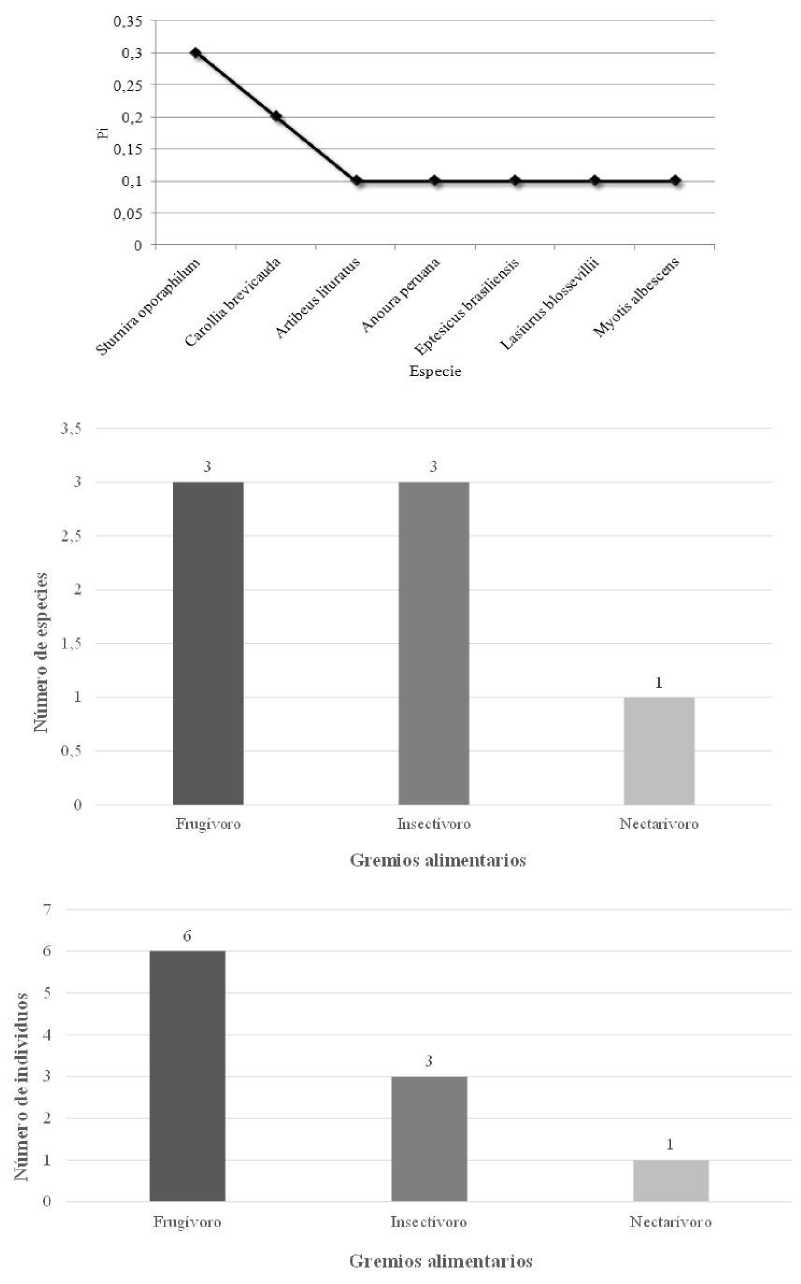
Datos de la comunidad de quirópteros capturados en Pastizal rodeado de bosque nativo (S3) de la parroquia Simón Bolívar, cantón Gonzalo Pizarro, provincia de Sucumbíos, Ecuador. Arriba: Curva de dominancia-diversidad, Pi = Proporción de individuos. Centro: Riqueza de especies. Abajo: Abundancia.
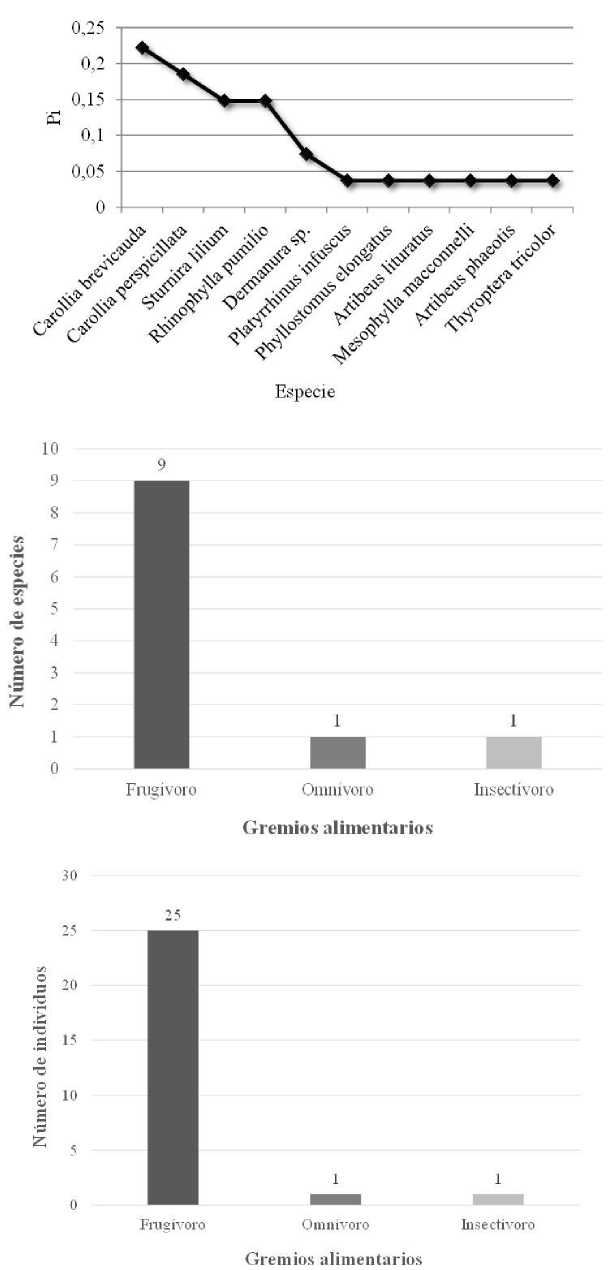
Datos de la comunidad de quirópteros capturados en Pastizal y chacra rodeados de bosque nativo (S4) de la parroquia Simón Bolívar, cantón Gonzalo Pizarro, provincia de Sucumbíos, Ecuador. Arriba: Curva de dominancia-diversidad, Pi = Proporción de individuos. Centro: Riqueza de especies. Abajo: Abundancia.
Estos pequeños espacios con poca cobertura vegetal seguramente no mantienen una comunidad de quirópteros establecida. Los murciélagos capturados seguramente fueron aquellos que provenían de los alrededores boscosos (donde crecen especies pioneras de arbustos y árboles como Piper sp. y Cecropia sp, cuyos frutos constituyen alimento importante para los murciélagos frugívoros).
La fauna de quirópteros en el área estudiada es diversa y abundante, con seguridad el muestreo en otros lugares de la gradiente del área incrementarán la riqueza. Los murciélagos presentes en la zona de estudio corresponden en su mayoría a especies de bosques húmedos tropicales bajos, aunque capturamos también una especie de distribución montana (Anoura peruana). La riqueza, abundancia, diversidad, así como la composición trófica varía entre los sitios estudiados. La mayor riqueza y abundancia la representan los murciélagos frugívoros (Phyllostominae y Stenodermatinae) dispersores de semillas. En todos los sitios los frugívoros fueron los más abundantes, las especies más abundantes en todos los sitios fueron los frugívoros de la subfamilia Carollinae; aunque otro grupo abundante fueron los nectarívoros Glossophaginae presentes en tres de los cuatro sitios.
Las especies dominantes en los cuatro sitios estudiados fueron diferentes, pero tres de los sitios fueron dominados por especies fugívoras. Los patrones de abundancia de frugívoros en el área de estudio es similar a los patrones encontrados en otros lugres del Neotrópico [30, 31], lo cual se explicaría por la permanente oferta de frutos [31, 32]. Este estudio muestra la gran diversidad de murciélagos de la familia Phyllostomidae endémicos de la región Neotropical [9, 33]. El 83 % de las especies y el 94,53 % de los individuos registrados pertenecen a esta familia. Estos murciélagos juegan roles ecosistémicos importantes al dispersar las semillas, polinizar las plantas y alimentarse de invertebrados y vertebrados [15, 34, 35].

Dendrogramas mostrando el análisis de similitud (índice Jaccard) de especies (arriba) y grupos tróficos (abajo) de la comunidad de quirópteros en cuatro sitios en la parroquia Simón Bolívar, cantón Gonzalo Pizarro, provincia de Sucumbíos, Ecuador, entre el 16 y 26 de marzo de 2013.
Un agradecimiento especial al Ing. Luciano Cepeda y a la Dra. Judith Ayala por el apoyo y facilidades en el campo, a Mesías Silva y Edwin Rebolier, empleados del Proyecto Coca Codo Sinclair, por su valiosa colaboración y asistencia en el trabajo de campo.
Agradecimientos
Este estudio formó parte del proyecto Estudio Florísti- co y Faunístico del Área de la Vía de Acceso a la casa de máquinas del Proyecto Hidroeléctrico Coca Codo Sinclair, ejecutado por el Museo Ecuatoriano de Ciencias Naturales del Instituto Nacional de Biodiversidad. Un agradecimiento especial al Ing. Luciano Cepeda ya la Dra. Judith Ayala por el apoyo y facilidades en elcampo, a Mesías Silva y Edwin Rebolier, empleados delProyecto Coca Codo Sinclair, por su valiosa colabora-ción y asistencia en el trabajo de campo.
Referencias
Ter Steege, H., Pitman, N., Sabatier, D., Castellanos, D., Van der Hout, P., Daly, D., Silveira, M., Phillips, O., Vásquez, R., Van Andel, T., Duivenvoorden, J.F., Adalardo, A., Ek, R., Lilwah, R., Thomas, R., Van Essen, J., Baider, C., Maas, P., Mori, S., Terborgh, J., Núñez, P., Mogollón, H. & Morawetz, W. (2003). A spatial model of tree a diversity and tree density for the Amazon. Biodiversity and Conservation, 12 (11), 2255-2277. doi: http://doi.Org/10.1023/A:1024593414624
Vázquez, A. M. & Ulloa, R. (1997). Estrategia para la conservación de la diversidad biológica en el sector forestal del Ecuador. Quito: Proyecto FAO-PAFE y EcoCiencia.
Chase J. & Leibold, M. (2003). Ecological niches: Linking classical and contemporary approaches. Chicago: University of Chicago Press.
Tilman, D. & Pacala, S. (1993). The maintenance of species diversity in plant communities. En R. Ricklefs, & D. Schluter (Eds.), Species diversity in ecological communities (pp. 13-25). Chicago: University of Chicago Press.
Myers, N. (1988). Threatened biotas "hotspots” in tropical forest. The Environmentalist, 8, 1-20. doi: http://doi.org/10.1007/BF02240252
Valencia, R., Balslev, H. & Paz y Miño, G. (1994). High tree alpha-diversity in Amazonian Ecuador. Biodiversity and Conservation, 3 (1), 21-28. doi: http://doi.org/10.1007/BF00115330
Mittermeier, R.A., Mittermeier, C.G., Brooks, T.M., Pilgrim, J.D., Konstant, W.R., Da Fonseca, G.A.B. & Kormos, C. (2003). Wilderness and biodiversity conservation. Proceedings of the National Academy of Sciences, 100(8), 1030910313. doi: http://doi.org/10.1073/pnas.1732458100
Tirira, D. (2011). Libro rojo de los mamíferos del Ecuador. 2° edición. Publicación Especial sobre los mamíferos del Ecuador 8.Quito: Fundación Mamíferos y Conservación, Pontificia Universidad Católica del Ecuador y Ministerio del Ambiente del Ecuador.
Tirira, D. (2014). Mamíferos del Ecuador: lista actualizada de especies/Mammals of Ecuador: Updapted checklist species. Versión 2014.2. Quito: Asociación Ecuatoriana de Mas- tozoología y Fundación Mamíferos y Conservación. URL: http://www.mamiferosdelecuador.com
Fleming, T.H. & Heithaus, E.R. (1981). Frugivorous bats, seed shadows, and the structure of the tropical forest. Biotropica, 13(2), 45-50. doi: http://doi.org/10.2307/2388069
Howe, H. F. & Smallwood, J. (1982). Ecology of seed dispersal. Annual Review of Ecology and Systematics, 13, 201228. doi: http://doi.org/10.1146/annurev.es.13.110182.001221
Connell, H. & Slatyer, R. (1977). Mechanisms of succession in natural communities and their role in community stability and organization. The American Naturalist, 111(982), 1119-1144. doi: http://doi.org/10.1086/283241
Guevara, S., Meave, J., Moreno, P.C., Laborde, J. & Castillo, S. (1994). Vegetación y flora de potreros en la sierra de los Tuxtlas, México. Acta Botánica Mexicana, 28, 1-27. doi: http://doi.org/10.21829/abm28.1994.714
Medellín, R. & Gaona, O. (1999). Seed dispersal by bats and birds in forest and disturbed habitats of Chiapas, México. Biotropica, 31(3), 478-485. doi: http://doi.org/10.1111/j.1744-7429.1999.tb00390.x
Kalko, E. (1998). Organization and diversity of tropical bat communities through space and time. Zoology, 101, 281297.
Kalko, E., Handley, C. & Handley D. (1996). Organization, diversity and long-term dynamics of a neotropical bat community. En M. L. Cody, & J. A. Smallwood (Eds.) LongTerm Studies of Vertebrate Communities (pp. 503-553). San Diego: Academic Press. doi: https://dx.doi.org/10.1016/b978-012178075-3/50017-9
Soriano, P. (2000). Functional structure of bat communities in tropical rainforests and Andean cloud forests. Ecotropicos, 13(1), 1-20. URL: http://webdelprofesor.ula.ve/ciencias/pascual/Pascual-PDFs/Ecotropicos-00a.pdf
Flores-Saldaña, M. (2008). Estructura de las comunidades de murciélagos en un gradiente ambiental en la reserva de la biosfera y tierra comunitaria de origen Pilón Pajas, Bolivia. Mastozoología Neotropical, 15(2), 309-322. URL: http://ref.scielo.org/gr4gkh
Palacios, W., Cerón, C., Valencia, R. & Sierra, R. (1999). Las formaciones Naturales de la Amazonía del Ecuador. En R. Sierra (Ed.), Propuesta preliminar de un Sistema de Clasificación de Vegetación para el Ecuador Continental. Quito: Proyecto INEFAN/GEF-BIRF y EcoCiencia.
Sierra, R. (1999). Propuesta preliminar de un Sistema de Clasificación de Vegetación para el Ecuador Continental. Quito: Proyecto GEF/INEFAN-BIRF y EcoCiencia.
Suárez, L. & Mena, P. (1994). Manual de métodos para inventarios de vertebrados terrestres. Quito: EcoCiencia.
Albuja, L. (1999). Murciélagos del Ecuador. 2da edición. Quito: Cicetrónic Cia. Ltda. Offset.
Tirira, D. (2007). Guía de campo de los Mamíferos del Ecuador. 2da edición. Quito: Ediciones Murciélago Blanco.
Magurran, A.E. (1988). Ecological diversity and its measurement. New Jersey: Princeton University Press.
Jost, L. (2006). Entropy and diversity. Oikos, 113(2), 363375. doi: http://doi.org/10.1111/j.2006.0030-1299.14714.x
Jost, L. (2007). Partitioning diversity into independent alpha and beta components. Ecology, 88(10), 2427-2439. doi: http://doi.org/10.1890/06-1736.1
Hair, J.F., Anderson, R. E., Tatham, R. L. & Black, W. C. (1999). Análisis Multivariante. 5a edición. Madrid: Prentice Hall Iberia.
UICN. (2013). IUCN Red List of Threatened Species. Versión 2012.2. URL: www.iucnredlist.org
Coca Codo Sinclair. (2013). Flora y fauna representativa de los bosque piemontanos y montano bajo del Proyecto Hidroeléctrico Coca Codo Sinclair. Publicación Técnico-Divulgativa de la Empresa Pública Estratégica Hidroeléctrica Coca Codo Sinclair. Quito: Imprenta Murgráfica.
Bolaños, T. (2000). Dieta y estructura trófica de la comunidad de Chiroptera en cuatro áreas de bosque andino en el sector sur-occidental de la sabana de Bogotá, Colombia. Trabajo de grado. Departamento de Biología. Facultad de Ciencias Pontificia Universidad Javeriana. Bogotá, Colombia.
Soriano, P. (1983). La comunidad de quirópteros de las selvas nubladas de los Andes de Mérida. Patrón reproductivo de los murciélagos frugívoros y las estrategias fenológicas de las plantas. Tesis de Maestría,Universidad de los Andes, Mérida.
Rivas, P., Sánchez, P. & Cadena, A. (1996). Estructura trófica de la comunidad de quirópteros en bosques de galería de la Serranía de la Macarena (Meta, Colombia). En H. H. Ge- noways, & R. J. Baker (Eds.), Contributions in Mammalogy: A memorial volume honoring Dr. J. Knox Jones, Jr. (pp. 237248). Lubbock: Texas Press Tech.
Mantilla-Meluk, H., Jiménez-Ortega, A.M., & Baker, R.J. (2009). Phyllostomid bats of Colombia: Annotated checklist, distribution, and biogeography. Special Publications Museum of Texas Tech University, 56, 1-37. URL: http://www.nsrl.ttu. edu/publications/specpubs/sps/SP56.pdf
Timm, R. (1994). The mammal fauna. En L. McDade, K. Bawa, H. Hespenheide, & G. Hartshorn (Eds.), La Selva: ecology and natural history of a Neotropical rainforest (pp. 229-237). Chicago: University of Chicago Press.
Patterson, B., Willig, M., & Stevens, R. (2003). Trophic strategies, niche partitioning, and patterns of ecological organization. En T. Kunz y M. Fenton (Eds.). Bat ecology (pp. 536-579). Chicago: University of Chicago Press. URL: http://hydrodictyon.eeb.uconn.edu/people/willig/Willig_pdf/SJ_109Patterson_etal_2003.pdf
Notas