Ciencias de la vida
Mejoramiento de la tolerancia a estrés hídrico en maíz en un programa de mejoramiento público
Mejoramiento de la tolerancia a estrés hídrico en maíz en un programa de mejoramiento público
Avances en Ciencias e Ingenierías, vol. 16, núm. 2, e3305, 2024
Universidad San Francisco de Quito

Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial 4.0 Internacional.
Recepción: 01 Mayo 2024
Aprobación: 30 Junio 2024
Resumen: A nivel global, gran parte de la producción de maíz se realiza bajo condiciones de secano, en las que la principal limitante para el rendimiento suele ser la disponibilidad hídrica. En maíz, el intervalo entre la floración masculina o antesis y la floración femenina o silking (ASI, en días) ha resultado ser un rasgo secundario útil dada su alta correlación con un mejor rendimiento en grano en ambientes propensos al estrés hídrico, su alta heredabilidad y la posibilidad de medirlo de manera rápida y precisa a campo. El objetivo de este trabajo fue caracterizar y seleccionar un conjunto de líneas endocriadas por su tolerancia al estrés hídrico. Para ello, se evaluaron a campo 240 líneas pertenecientes al Programa de Mejoramiento de Maíz Templado del INTA, en Pergamino durante la campaña 2019/20 bajo condiciones de alta densidad de siembra (16 pl.m-2) en un diseño de bloques completos al azar con 2 repeticiones. Un subconjunto de 50 líneas se evaluó nuevamente en el mismo sitio en la campaña 2020/21 (correspondiente a la fase La Niña del fenómeno climático El Niño Oscilación del Sur - ENOS). Las líneas se caracterizaron por ASI y rasgos agronómicos, como el porcentaje de vuelco y de quebrado. La alta densidad poblacional promovió la expresión de diferencias genotípicas significativas en ASI (p < 0,05, media: 2,3 días, rango: -3 a +9 días), porcentajes de vuelco (p < 0,05, media: 26,5%, rango: 0 a 100%) y quebrado (p < 0,05, media: 5,3%, rango: 0 a 80%). Los valores de heredabilidad fueron de 63, 51 y 64, para ASI, % de vuelco y % de quebrado, respectivamente. En 2020/21, se evaluaron las 50 líneas que el año previo había registrado los menores valores de ASI (media = 0,52), y que a su vez tuvieron porcentajes de vuelco menor al 20% y de quebrado de tallo menor al 5%, encontrando diferencias significativas entre líneas (para ASI: p < 0,05, media: 1,8, rango: -3 a +6 días). En base a los datos obtenidos en ambos años, se seleccionó un conjunto de líneas representativas de los diferentes grupos heteróticos del programa de mejoramiento para realizar cruzamientos biparentales y desarrollar nuevas líneas de menor ASI y, por ende, mejor comportamiento frente a situaciones de estrés. Para ello, la endocría a partir de la generación S0 y la evaluación de las líneas segregantes en combinación híbrida se realizarán bajo alta densidad de siembra (16 pl.m-2). Este método de selección permitirá el desarrollo de germoplasma tolerante al estrés.
Palabras clave: sequía, estabilidad del rendimiento, líneas endocriadas, eficiencia de uso de recursos, rasgos secundarios, alta densidad poblacional.
Abstract: World maize production is mainly carried out in rainfed systems in which the main limitation to grain yield is usually water availability. In maize, the anthesis-silking interval (ASI, in days) is proved to be a useful secondary trait as it is highly correlated with improved grain yield in drought-prone environments, has high heritability and can be fast and accurately measured in the field. The objective of this work was to characterize and select a set of inbred lines for their tolerance to water stress. To achieve this, 240 inbred lines of the INTA Pergamino Temperate Maize Breeding Program were grown in the field during the 2019-20 season at high planting density (16 pl.m-2) in a randomized complete block design with 2 replicates. A subset of 50 inbreds was sown in the same site during the dry 2020-21 growing season. Inbreds were characterized for ASI and defensive traits, such as root and stalk lodging. High stand density promoted the expression of significant genotypic differences in ASI (p < 0.05, mean: 2.3 d, range: -3 to +9 d), root (p < 0.05, mean: 26.5%, range: 0 to 100%) and stalk lodging (p < 0.05, mean: 5.3%, range: 0 to 80%). Heritabilities values were 63, 51 and 64 for ASI, root and stalk lodging, respectively. In 2020-21, the 50 inbreds with the lowest ASI values (mean = 0.52), and with less than 20% and 5% root and stalk lodging, respectively were evaluated again, finding significant differences between inbreds (for ASI: p<0.05, mean: 1.8, range: -3 to +6 days). Based on the data obtained in both years, a set of inbreds representative of the different heterotic groups of the breeding program was selected to make biparental crosses and develop new inbreds with shorter ASI, and therefore, with better performance under stressful conditions. For this, the inbreeding from the S0 generation and the evaluation of inbreds in hybrid combination will be carried out at high planting density (16 pl.m-2). This selection method will allow the development of stress-tolerant germplasm.
Keywords: drought, yield stability, inbreds, resource use efficiency, secondary traits, high plant density.
INTRODUCCIÓN
En el cultivo de maíz (Zea mays L.), el rendimiento en grano está principalmente determinado por el número de granos por planta [1, 2] que se fija durante el periodo crítico de 30 días alrededor de la fase de floración [3, 4]. El número de granos fijados está asociado a: (i) las tasas de crecimiento de la espiga y de la planta durante el periodo crítico [5]; (ii) la partición de biomasa a la espiga durante dicho periodo [6, 7]; (iii) la sincronía entre las floraciones masculina (antesis) y femenina (silking) [8], que determinan el intervalo antesis-silking (ASI, por su sigla en inglés); y (iv) la sincronía en la polinización de flores dentro de la espiga y entre espigas de una misma planta [9], que determina la prolificidad (espigas con granos por planta).
El maíz se cultiva en más de 200 millones de hectáreas a nivel global [10], lo que lo expone a una variedad de condiciones climáticas y a múltiples estreses bióticos y abióticos. Buena parte de esa superficie se encuentra bajo condiciones de secano, en las que la principal limitante para el rendimiento es la disponibilidad hídrica [11]. El maíz es sensible al estrés hídrico durante la mayor parte de los estadios, pero su susceptibilidad es mayor en el periodo alrededor de la floración [12, 13]. Al ocurrir un déficit hídrico en la floración, se produce un retraso en el crecimiento de la espiga y, en consecuencia, en la aparición de los estigmas respecto a la panoja y a la emisión de polen [14, 15].
El uso de caracteres secundarios cuya varianza genética y heredabilidad son elevadas bajo estrés, resulta útil para aumentar la eficiencia de selección [16, 17, 18]. El éxito de la mejora del rendimiento mediante selección indirecta por un rasgo secundario dependerá de encontrar, al mismo tiempo, una alta correlación entre los efectos aditivos que determinan a ambos caracteres junto con una elevada heredabilidad y facilidad en su medición [19, 20]. El ASI es un rasgo ampliamente utilizado por mejoradores y fisiólogos para describir y seleccionar la tolerancia al estrés hídrico en el maíz [4, 21]. Trabajos realizados con maíces tropicales sometidos al estrés hídrico en floración muestran la alta correlación del rendimiento con el ASI y la prolificidad. Estos rasgos, a su vez, bajo condiciones de estrés severo, registraron valores de heredabilidad superiores a los del rendimiento en grano [17]. Además, el ASI, la prolificidad y la senescencia foliar, mostraron valores altos de heredabilidad en ambientes deficitarios en N, y su uso junto al rendimiento mejoró la eficiencia de selección en dichos ambientes [18].
El estrés por densidad poblacional puede considerarse un estrés crónico producto de la exposición en simultáneo a múltiples factores ambientales que reducen la captura o utilización de recursos de cada planta individual [22, 23]. Los factores ambientales que componen el estrés son difíciles de distinguir, pero se los puede considerar como desvíos de las condiciones óptimas de crecimiento, que limitan a que la planta alcance su potencial genético. La alta densidad poblacional es un estrés fácil de gestionar en su intensidad, adecuado para la evaluación de un gran número de genotipos, afecta a las plantas durante todo su ciclo y el rango de niveles de estrés que permiten discernir el comportamiento de los genotipos es amplio [24]. Los programas de mejoramiento comercial utilizan altas densidades poblacionales para evaluar el germoplasma, pero esa evaluación se realiza en fases avanzadas en el desarrollo de híbridos. Para aumentar la tasa de ganancia genética, la selección de un rasgo en la población segregante resulta superior frente a la selección en estados avanzados, cuando el número de genotipos selectos es acotado. Algunos autores proponen incorporar evaluaciones en alta densidad en etapas tempranas del desarrollo de líneas endocriadas, para mejorar la tolerancia al estrés [24].
El objetivo de este trabajo fue caracterizar y seleccionar, por su tolerancia al estrés hídrico, un conjunto de líneas endocriadas representativas del germoplasma disponible en el Programa de Mejoramiento de Maíz Templado del INTA. A partir de dicha selección se espera iniciar un programa de desarrollo de líneas endocriadas con mayor tolerancia al estrés, particularmente hídrico, a partir de la combinación de dos herramientas: la selección temprana bajo condiciones de alta densidad y el uso de un atributo secundario altamente correlacionado con la tolerancia al estrés hídrico, como el ASI. Para iniciar este programa, se caracterizaron 240 líneas endocriadas en condiciones de estrés por alta densidad durante dos campañas. Luego se seleccionaron líneas que dieron origen a poblaciones segregantes producto de cruzamientos biparentales entre líneas de desempeño superior.
MATERIALES Y MÉTODOS
En la campaña agrícola 2019/2020 (año 1) se realizó la evaluación a campo de 240 líneas endocriadas pertenecientes al Programa de Mejoramiento de Maíz Templado del INTA. Estas líneas representan la variabilidad genética disponible en el programa. En la campaña 2020/2021 (año 2) se evaluaron 50 líneas seleccionadas a partir del análisis realizado en el año 1. Ambos experimentos se llevaron adelante en el campo experimental de la EEA Pergamino del INTA.
Las siembras se realizaron el 29 de octubre de 2019 y el 2 de noviembre de 2020 en un suelo argiudol típico serie Pergamino bajo siembra directa, con un diseño en bloques completos al azar con 2 repeticiones. La densidad de siembra utilizada fue de 16 pl.m-2, el doble de la usualmente empleada. A la siembra se fertilizó con 200 kg.ha-1 de urea y 150 kg.ha-1 de mezcla arrancadora (7N 40P 5S) y los experimentos se condujeron en secano. La unidad experimental consistió en parcelas de 5 metros de largo por 2 surcos de ancho, distanciados a 0,7 m. En cada parcela se registraron las fechas en las que el 50 % de las plantas de los 4 m centrales de ambos surcos alcanzaron antesis y silking. A partir de la diferencia entre ambas fechas se estimó el ASI en días. Además, se cuantificaron características agronómicas como los porcentajes de plantas con vuelco de raíz y con quebrado de tallo. Los datos fueron estudiados mediante el análisis de la varianza. A partir de los cuadrados medios se estimaron los componentes de la varianza y, en base a estos, la heredabilidad.
RESULTADOS
El promedio de las temperaturas medias diarias durante el ciclo del cultivo fue de 22,1 °C y 21,7 °C para el año 1 y 2, respectivamente (Fig. 1), en tanto que las precipitaciones fueron de 665 mm y 398 mm cada año. Para el periodo crítico (i.e., 30 días centrados en la floración), el promedio de temperaturas medias fue de 22,8 °C y 22,7 °C, respectivamente, y las precipitaciones fueron de 89 mm y 144 mm cada año. El año 1 se caracterizó por la ocurrencia de 3 días de golpe de calor (i.e., días con temperatura máxima por encima de los 35 °C) en las semanas previas al inicio del periodo crítico (Fig. 1), lo cual ocasionó quemado foliar en unos pocos genotipos. El año 2 se caracterizó por escasas precipitaciones y fue caracterizado como correspondiente a la fase La Niña del fenómeno El Niño Oscilación del Sur (ENOS), que para la región del sitio experimental significa lluvias por debajo del promedio.
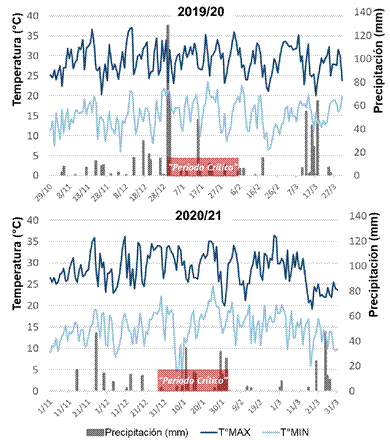
Fig. 1
Condiciones climáticas exploradas en las campañas en que se realizó la caracterización del germoplasma. T°MAX: temperatura máxima diaria, T°MIN: temperatura mínima diaria. El rectángulo rojo indica el lapso en el que ocurrió el periodo crítico como promedio de todo el germoplasma evaluado en cada año.
Figura 1. Condiciones climáticas exploradas en las campañas en que se realizó la caracterización del germoplasma. T°MAX: temperatura máxima diaria, T°MIN: temperatura mínima diaria. El rectángulo rojo indica el lapso en el que ocurrió el periodo crítico como promedio de todo el germoplasma evaluado en cada año.
En ambos años, los genotipos evaluados mostraron diferencias significativas en los valores de ASI, porcentaje de vuelco y porcentaje de quebrado (p < 0,05). El primer año se caracterizaron 240 líneas, con valores promedio de 2,3 días para el ASI (rango: -3 a 9 días); 26,5 % para vuelco (rango: 0 a 100 %); y 5,3 % para quebrado (rango: 0 a 80,1 %) (Fig. 2). Los valores de heredabilidad fueron de 0,63; 0,51 y 0,64, para ASI, porcentaje de vuelco y porcentaje de quebrado, respectivamente. A partir de estos datos, se seleccionaron las 50 líneas con desempeño superior para estos rasgos (i.e., menores valores de ASI, y valores de porcentaje de vuelco y quebrado menores al 20 % y al 5, respectivamente). El valor medio de ASI en el año 1 de las 50 líneas selectas fue de 0,52 días (Fig. 3). En el año 2, los valores promedio observados fueron 1,8 días para el ASI (rango: -3 a 6 días); 19,6% para vuelco (rango: 0 a 65 %); y 45,9 % para quebrado (rango: 0 a 100 %). Esta información permitió construir un ranking de las líneas de acuerdo con el desempeño que tuvieron para estas características.
El ranking de genotipos permitió identificar líneas de buen desempeño dentro de cada grupo heterótico u origen genético, y así dar inicio al desarrollo de nuevas líneas con comportamiento superior. En la campaña 2021/2022 se realizaron 8 cruzas biparentales. En 2022/2023, a partir de la autofecundación de individuos F1 de cada cruza, se originó la semilla S0 (F2). En 2023/2024 se sembraron las poblaciones S0 en condiciones de alta densidad para continuar con el proceso de endocría. En las campañas próximas se espera continuar con la endocría bajo siembras de alta densidad. A partir de la generación S2 se iniciarán los cruzamientos por probadores de los diferentes grupos heteróticos, para iniciar la selección por aptitud combinatoria, tanto en condiciones de densidad de siembra normal como de alta densidad.
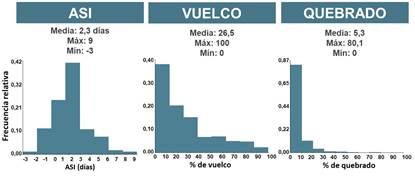
Fig. 2
Diagramas de frecuencias relativas observadas en 240 líneas evaluadas en el año 1 para los caracteres ASI, porcentaje de vuelco y porcentaje de quebrado.
Figura 2. Diagramas de frecuencias relativas observadas en 240 líneas evaluadas en el año 1 para los caracteres ASI, porcentaje de vuelco y porcentaje de quebrado.
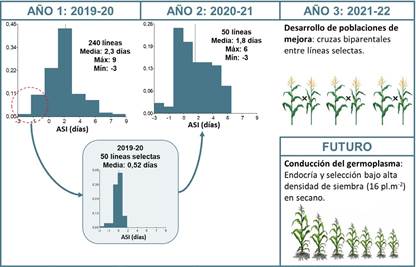
Fig. 3
Esquema de mejoramiento propuesto.
Figura 3. Esquema de mejoramiento propuesto.
DISCUSIÓN
El proceso de mejoramiento para la obtención de híbridos requiere de numerosas generaciones de endocría. A lo largo de más de 100 años de historia en el desarrollo de híbridos de maíz [25, 26, 27] ocurrió una selección indirecta que produjo una reducción del ASI [28, 29, 30]. Aun así, bajo condiciones de estrés, se encontraron diferencias en el ASI de los materiales evaluados, indicando que existe aún variabilidad genética para este rasgo, al menos para el germoplasma del Programa de Mejoramiento de Maíz Templado del INTA. Al tratarse de un programa de mejoramiento público, en el cual: (i) la intensidad de selección no ha sido tan elevada como en los programas comerciales, (ii) la variabilidad genética disponible es mucho mayor, y (iii) se incorporan nuevas fuentes de variabilidad genética frecuentemente, es posible que pueda haber una mayor respuesta a la selección que aquella que podría obtenerse en programas comerciales.
De acuerdo con nuestro conocimiento, los programas de mejoramiento comercial realizan selección en altas densidades de siembra (11–16 pl.m-2), a través de la evaluación del comportamiento de combinaciones híbridas. Esta selección ocurre generalmente en estados avanzados y sin someter a estrés a las líneas en desarrollo per se. Trabajos previos han demostrado que las líneas endocriadas podrían resultar incluso más susceptibles al estrés hídrico que los híbridos, dada la mejor adaptación que les confiere, a los segundos, la heterosis que expresan [31, 32, 33]. En este trabajo proponemos iniciar la selección en condiciones de alta densidad desde el desarrollo de las líneas, conduciendo el vivero de cría en alta densidad.
Al realizar la endocría en estas condiciones, en primer lugar se asegura la ocurrencia de estrés, sin necesidad de realizar evaluaciones fuera del ambiente objetivo del programa de mejora y/o con experimentos manipulativos que impidan la infiltración de las precipitaciones, difíciles de conducir en grandes extensiones a campo. Segundo, indirectamente, a través del proceso de autofecundación, se seleccionan aquellos individuos con menor ASI bajo estrés. En los individuos que presenten mayor susceptibilidad al estrés, la floración femenina tendrá un desfase respecto a la antesis que no permitirá la autofecundación exitosa, aumentando la intensidad de selección por este rasgo.
A su vez, no debe perderse de vista el rendimiento potencial, por lo cual, se propone realizar los ensayos de topcross (i.e., evaluación en combinación híbrida de las líneas en desarrollo para su selección por habilidad combinatoria) no solo bajo condiciones de alta densidad, sino también bajo las densidades de siembra habitualmente empleadas y en múltiples localidades [34]. En aquellos programas de mejoramiento que han integrado el uso de haploides duplicados para el desarrollo de líneas, se propone el fenotipado de estos rasgos en el primer año de evaluación. Finalmente, en programas que además integren selección genómica, es importante incorporar a los modelos de selección este tipo de rasgos de alta heredabilidad y fuerte correlación con la estabilidad del rendimiento.
CONCLUSIONES
La integración de conocimientos de diferentes disciplinas como la genética cuantitativa, la ecofisiología, la estadística, la genética molecular, entre otras, continúa siendo crítica para el desarrollo de nuevo germoplasma con mayor rendimiento y estabilidad. Este trabajo ha permitido identificar fuentes de variación útiles para el desarrollo de germoplasma con mayor tolerancia al estrés abiótico, en particular hídrico. A su vez, utilizando el esquema de selección aquí propuesto (Fig. 3) se ha logrado establecer un programa de mejoramiento de la tolerancia al estrés que permitirá el desarrollo de germoplasma de maíz templado adaptado a la región pampeana argentina.
En los años siguientes, empleando el conjunto de 240 líneas aquí evaluadas, continuarán los estudios de las bases genéticas de rasgos asociados con la determinación del rendimiento y su estabilidad. Los caracteres a estudiar serán: el número de flores diferenciadas en la floración, la velocidad de emisión de estigmas, el número de hileras, la relación entre el número de flores diferenciadas y el número de granos fijados, entre otros. Además de profundizar en el entendimiento de los procesos fisiológicos relacionados con la tolerancia al estrés, esto permitirá continuar con el desarrollo de germoplasma y el enriquecimiento de poblaciones de mejora con alelos que incrementen su adaptación a situaciones de estrés. A futuro, se espera poder establecer un programa de selección recurrente que pueda proveer fuentes de variación valiosas a los programas de mejoramiento de maíz de Argentina.
AGRADECIMIENTOS
Este trabajo fue financiado por los proyectos de INTA cartera 2019: I-128 y cartera 2023: I110 e I102, y por el convenio de vinculación tecnológica INTA Semilleros. Las autoras desean agradecer a Macarena Mauer, Leonardo Magro, Mauro Nazar, Oscar Ale y Renzo Rodríguez por la ayuda técnica para la implantación y mantenimiento de los experimentos.
Agradecemos también a la Red Latinoamericana de Maíz y al proyecto Tech Maíz del Programa Iberoamericano de Ciencia y Tecnología para el Desarrollo (CYTED) por promover el intercambio y la gestión del conocimiento en la región.
CONTRIBUCIONES DE LOS AUTORES
María Otegui y Luciana Galizia concibieron la investigación y desarrollaron la metodología. Luciana Galizia y Fátima Palifermo realizaron el trabajo de campo. Luciana Galizia realizó el análisis de los datos. Luciana Galizia y María Otegui redactaron el manuscrito.
CONFLICTO DE INTERÉS
Las autoras declaran que no existe ningún conflicto de interés en la presente investigación.
Agradecimientos
Este trabajo fue financiado por los proyectos de INTA cartera 2019: I-128 y cartera 2023: I110 e I102, y por el convenio de vinculación tecnológica INTA Semilleros. Las autoras desean agradecer a Macarena Mauer, Leonardo Magro, Mauro Nazar, Oscar Ale y Renzo Rodríguez por la ayuda técnica para la implantación y mantenimiento de los experimentos.
Agradecemos también a la Red Latinoamericana de Maíz y al proyecto Tech Maíz del Programa Iberoamericano de Ciencia y Tecnología para el Desarrollo (CYTED) por promover el intercambio y la gestión del conocimiento en la región.
REFERENCIAS
[1] Cirilo, A.G. y Andrade, F.H. (1994). Sowing date and maize productivity: II. Kernel number determination. Crop Science, 34, 1044-1046. doi: https://doi.org/10.2135/cropsci1994.0011183X003400040038x
[2] Otegui, M.E. (1995). Prolificacy and grain yield components in modern Argentinean maize hybrids. Maydica, 40, 371-376. https://www.researchgate.net/publication/261473724_Prolificacy_and_Grain_Yield_Components_in_ Modern_Argentinian_Maize_Hybrids
[3] Fischer K.S. y Palmer, A.F.E. (1984). Tropical Maize. In: P.R. Goldsworthy y N.M. Fisher (Eds.), The physiology of tropical field crops (pp. 213-248). J. Wiley & Sons Ltd. https://books.google.com.ec/books/about/The_Physiology_of_ Tropical_Field_Crops.html?id=4fTwAAAAMAAJ&redir_esc=y
[4] Borrás, L. y Vitantonio-Mazzini, L.N. (2018). Maize reproductive development and kernel set under limited plant growth environments. Journal of Experimental Botany, 69(13), 3235–3243. doi: https://doi.org/10.1093/jxb/erx452
[5] Andrade, F.H., Vega, C., Uhart, S., Cirilo, A.G., Cantarero, M. y Valentinuz, O. (1999). Kernel number determination in maize. Crop Science, 39, 453-459. https://www.semanticscholar.org/paper/Kernel-Number-Determination-in-MaizeAndrade-Vega/d3c00d4a8b827b0b52826600948d1c61808fa354
[6] Pagano, E. y Maddonni, G.A. (2007). Intra-specific competition in maize: early established hierarchies differ in plant growth and biomass partitioning to the ear around silking. Field Crops Research, 101, 306-320. doi: https://doi. org/10.1016/j.fcr.2006.12.007
[7] Borrás, L., Westgate, M.E., Astini, J.P. y Echarte, L. (2007). Coupling time to silking with plant growth rate in maize. Field Crops Research, 102, 73-85. doi: https://doi.org/10.1016/j.fcr.2007.02.003
[8] Bolaños, J. y Edmeades, G.O. (1993). Eight cycles of selection for drought tolerance in lowland tropical maize. II. Responses in reproductive behavior. Field Crops Research, 31, 253-268. doi: https://doi.org/10.1016/03784290(93)90065-U
[9] Cárcova, J., Uribelarrea, M., Borrás, L., Otegui, M. E. y Westgate, M.E. (2000). Synchronous Pollination within and between Ears Improves Kernel Set in Maize. Crop Science, 40, 1056-1061. doi: https://doi.org/10.2135/ cropsci2000.4041056x
[10] FAO. (2024). Food and Agriculture Organization of the United Nations – Statistics Division. FAOSTAT. http://fao.org/ faostat/es/
[11] Lobell, D.B., Cassman, K.G. y Field, C.B. (2009). Crop yield gaps: their importance, magnitudes, and causes. Annual Review of Environment and Resources, 34, 179–204. doi: https://doi.org/10.1146/annurev.environ.041008.093740
[12] Hall, A.J., Lemcoff, J.H. y Trapani, N. (1981). Water stress before and during flowering in maize and its effects on yield, its components, and their determinants. Maydica, 26, 19–38. https://www.semanticscholar.org/paper/Water-stressbefore-and-during-flowering-in-maize-Hall-Lemcoff/9e1e0ebb66abaaf7705a52a7ab485cdf201406aa
[13] Grant, R.F., Jackson, B.S., Kiniry, J.R. y Arkin, G.F. (1989). Water deficit timing effects on yield components in maize. Agronomy Journal, 81, 61–65. doi: https://doi.org/10.2134/agronj1989.00021962008100010011x
[14] Herrero, M.P. y Johnson, R.R. (1981). Drought stress and its effects on maize reproductive systems. Crop Science, 21(1), 105–110. doi: https://doi.org/10.2135/cropsci1981.0011183X002100010029x
[15] Otegui, M.E., Andrade, F.H. y Suero, E.E. (1995). Growth, water use, and kernel abortion of maize subjected to drought at silking. Field Crops Research, 40, 87-94. doi: https://doi.org/10.1016/0378-4290(94)00093-R
[16] Ludlow, M.M. y Muchow, R.C. (1990). A critical evaluation of traits for improving crop yields in water-limited environments. Advances in Agronomy, 43, 107-153. doi: https://doi.org/10.1016/S0065-2113(08)60477-0
[17] Bolaños, J. y Edmeades, G.O. (1996). The importance of the anthesis silking interval in breeding for drought tolerance in tropical maize. Field Crops Research, 48, 65-80. doi: https://doi.org/10.1016/0378-4290(96)00036-6
[18] Bänziger, M. y Lafitte, H.R. (1997). Breeding tropical maize for low N environments. II. The values of secondary traits for improving selection gains under low N. Crop Science 37, 1110–1117.
[19] Blum, A. (1988). Plant Breeding for Stress Environments. CRC Press Inc. doi: https://doi.org/10.1201/9781351075718
[20] Falconer, D.S. y Mackay, T.F.C. (1996). Introduction to quantitative genetics. 4th ed. Pearson Education Limited. https:// archive.org/details/IntroductionToQuantitativeGenetics/mode/2up
[21] Rebolloza-Hernández, H., Cervantes-Adame, Y.F., Broa-Rojas, E., Bahena-Delgado, G. y Olvera-Velona, A. (2020). Fenotipeo y selección de líneas S1 segregantes de maíz tolerantes a estrés hídrico. Biotecnia XXII, (3), 20-28. doi: https://doi.org/10.18633/biotecnia.v22i3.1130
[22] Tollenaar, M. y Lee, E.A. (2002). Yield potential yield, yield stability and stress tolerance in maize. Field Crops Research, 75, 161–170. doi: https://doi.org/10.1016/S0378-4290(02)00024-2
[23] Mansfield, B.D. y Mumm, R.H. (2014). Survey of Plant Density Tolerance in U.S. Maize Germplasm. Crop Science, 54, 157-173. doi: https://doi.org/10.2135/cropsci2013.04.0252
[24] Lee, E. y Tollenaar, M. (2007). Physiological basis of successful breeding strategies for maize grain yield. Crop Science, 47, S202-S215. doi: https://doi.org/10.2135/cropsci2007.04.0010IPBS
[25] Shull, G.H. (1908). The composition of a field of maize. Journal of Heredity, 4, 296–301. doi: https://doi.org/10.1093/ jhered/os-4.1.296
[26] Shull, G.H. (1909). A pureline method of corn breeding. Journal of Heredity, 5, 51–59. doi: https://doi.org/10.1093/ jhered/os-5.1.51
[27] Andorf, C., Beavis, W.D., Hufford, M., Smith, S., Suza, W.P., Wang, K., Woodhouse, M., Yu, J. y Lübberstedt, T. (2019). Technological advances in maize breeding: past, present and future. Theoretical Applied Genetics, 132, 817–849. doi: https://doi.org/10.1007/s00122-019-03306-3
[28] Campos, H., Cooper, M., Edmeades, G. O., Löffler, C., Schussler, J. R., Ibañez, M. (2006). Changes in drought tolerance in maize associated with fifty years of breeding for yield in the U.S. Corn Belt. Maydica, 51: 369–381. researchgate.net/publication/271517539_Changes_in_drought_tolerance_in_maize_associated_with_fifty_ years_of_breeding_for_yield_in_the_US_Corn_Belt
[29] Welcker, C., Spencer, N. A., Turc, O., Granato, I., Chapuis, R., Madur, D., Beauchene, K., Gouesnard, B., Draye, X., Palaffre, C., Lorgeou, J., Melkior, S., Guillaume, C., Presterl, T., Murigneux, A., Wisser,R. J., Millet, E. J., van Eeuwijk, F., Charcosset, A. y Tardieu, F. (2022). Physiological adaptive traits are a potential allele reservoir for maize genetic progress under challenging conditions. Nature Communications, 13, 3225. https://doi.org/10.1038/s41467-022-30872-w
[30] dos Santos, C. L., Miguez, F. E., King, K. A., Ruiz, A., Sciarresi, C., Baum, M. E., Danalatos, G. J. N., Stallman, M., Wiley, E., Pico, L. O., Thies, A., Puntel, L. A., Topp, C. N., Trifunovic, S., Eudy, D., Mensah, C., Edwards, J. W., Schnable, P. S., Lamkey, K. R., Vyn, T.J. y Archontoulis, S. V. (2023). Accelerated leaf appearance and flowering in maize after four decades of commercial breeding. Crop Science, 63, 2750–2762. https://doi.org/10.1002/csc2.21044
[31] Echarte, L. y Tollenaar, M. (2006). Kernel set in maize hybrids and their inbred lines exposed to stress. Crop Science, 46, 870-878. doi: https://doi.org/10.2135/cropsci2005.0204
[32] Liu, W. y Tollenaar, M. (2009). Response of yield heterosis to increasing plant density in maize. Crop Science, 49, 18071816. doi: https://doi.org/10.2135/cropsci2008.07.0422
[33] Araus, J.L., Sánchez, C. y Cabrera-Bosquet, L. (2010). Is heterosis in maize mediated through better water use? New Phytologist, 187, 392-406. doi: https://doi.org/10.1111/j.1469-8137.2010.03276.x
[34] Cooper, M., Messina, C., Podlich, D., Radu Totir, L., Baumgarten, A., Hausmann, N.J., Wright, D. y Graham, G. (2014). Predicting the future of plant breeding: complementing empirical evaluation with genetic prediction. Crop and Pasture Science, 65(4), 11-336. doi: https://doi.org/10.1071/CP14007